

Невесомость (космический полет) является уникальной окружающей средой, которая создает ряд физиологических неблагоприятных проблем разным системам и органам тела человека. Микрогравитация, с которой сталкивается человек во время космической полета, или в моделях, имитирующих невесомость, вызывает изменения в функциях скелетных мышц. Изменения в сократительных свойствах мышцы в результате воздействий микрогравитации крайне сложны и многочисленные исследования поддерживают эту концепцию. Потеря массы и «слабость» скелетных мышц, отмечаемая еще ранних космических миссий человека на кораблях типа «Gemini», «Союз» и «Skylab», остается важной проблемой. Так уже в ранних кратковременных полетах было обнаружено отрицательное влияние невесомости на нервно-мышечную функцию. После полетов продолжительностью 5-11 суток было обнаружено снижение силовых показателей мышц на 14-30 % [4, 8, 10, 32] и снижение объема мышцы [26], уменьшение капилляризации мышц и площади поперечного сечения волокон типа I и II) [18]. После 17-суточного полета с использованием изокинетической динамометрии не было обнаружено значительного снижения не только максимальной произвольной силы, но и электрически вызванного (частота 50 имп/с) максимального суставного момента (10 %), развиваемого мышцами-разгибателями стопы [47]. Вместе с тем было обнаружено снижение силы сокращения отдельных мышечных волокон типа I (на 21 %) и типа IIa (на 25 %) m. soleus [57], тогда как волокна этих типов m. gastrocnemius практически не были затронуты [26]. Полет протяженностью 28 суток на «Skylab-2» показал большее снижение суставного момента мышц бедра по сравнению с мышцами рук и мышцами-разгибателями, чем сгибателей мышцы [28]. Полет 59 суточный на «Skylab-3» показал еще большие различия между суставным моментом, развиваемым мышцами бедра (на 20 %) и рук (лишь на 2 %). Однако когда члены экипажей выполняли физические упражнения для мышц ног [28], то различие в величине суставного момента, развиваемого мышцами бедра до и после 84-суточного полета «Skylab-4», было значительно меньшим (лишь на 6 %).
Подобные функциональные тесты были выполнены и после продолжительных космических экспедиций (до 237 суток) на орбитальной станции «МИР» и было обнаружено снижение до 50 % максимального суставного момента мышц-разгибателей бедра, мышц-сгибателей и разгибателей стопы [2, 15, 16, 17, 40, 60]. Более того отмечалось существенное уменьшение тощей массы тела, несмотря на выполнение в космических полетах физических тренировок (в течение 1-2 часов) с определенным комплексом упражнений [3, 60]. Однако используемый комплекс физических упражнений во время космических полетов полностью не предотвращал снижение функций и работоспособности скелетных мышц [6, 24, 26, 55]. Снижение сократительных свойств в космическом полете может иметь отрицательное значение как на этапах завершения операторских задач при выполнении вне корабельной деятельности и/или на поверхности Марса, так и по возвращении на Землю.
Большая часть информации о влиянии разгрузки на функции мышц была получена в наземных модельных исследованиях [2, 4, 6, 14].
Сравнительных исследований степени снижения сократительных функций мышц после космического полета и наземных аналогов, моделирующих невесомость, было выполнено в небольшом числе [4, 55]. Мы изучили изменение сократительных свойств ТМГ с участием семи космонавтов до и после продолжительных космических полетов (> 120 суток) на станции «МИР» и восьми взрослых испытуемых-добровольцев, находящихся в условиях 120-суточного постельного режима в антиортостатическом положении [6].
До сих пор, из-за методологических трудностей, собственно-сократительные свойства скелетных мышц человека и их изменения под влиянием реальной невесомости и/или моделей, имитирующей ее, не были предметом систематических исследований. Внимание исследователей было сконцентрировано, главным образом, на изучении механических особенностей произвольных сокращений/движений мышц. В последнем случае характеристики функционирования нервно-мышечного аппарата в значительной мере, если не целиком, определяются свойствами, как самих мышц, в частности сократительными свойствами мышечных волокон, составляющих эти мышцы, так и характером центральной иннервации (моторной посылкой). Изучение удельного вклада каждого из этих факторов становится возможным лишь при сопоставлении параметров произвольного сокращения и сокращения той же мышцы, вызванного электрическим тетаническим раздражением. Механические параметры мышцы, зарегистрированные во время электрического раздражения двигательного нерва, иннервирующего мышцу, позволяют исследовать периферические изменения.
Работа, представленная здесь, является первым исследованием с количественной оценкой степени изменений функциональных свойств нервно-мышечного аппарата у космонавтов после продолжительной космической миссии. Исследовались механические ответы трехглавой мышцы голени (ТМГ), как постуральной и антигравитационной мышцы в условиях 1-G [20]. Краткие сообщения о результатах работы были ранее представлены [39].
Основная цель данной работы состояла в том, что используя отдельную мышцу, участвующую в произвольном и электрически вызванном сокращении, определить механизмы, лимитирующие и определяющие сократительные свойства мышц у человека после продолжительного космического полета. Поскольку ранее мы использовали идентичный методологический подход в исследовании влияний продолжительного постельного режима на функции мышц-разгибателей стопы (на примере трехглавой мышцы голени – ТМГ) [6, 38], то вторая цель работы состояла в том, чтобы сравнить и проверить эффективность наземного постельного режима, как модели космического полета. Результаты этих работ были ранее представлены [22, 27].
Материалы и методы исследования
Испытуемые
В исследовании участвовала группа (n = 7) космонавтов-мужчин, участников основных экипажей (ОЭ-18, 19, 22, 25, 26, 27) в полетах на орбитальной станции «МИР», возрастом между 43 и 54 лет (в среднем 45.1 ± 2.0), ростом – 167 и 182 см (в среднем 176.0 ± 2.3) и массой тела – 70 и 86 (в среднем 79.9 ± 2.0). Длительность космической миссии составляла 213.0 ± 30.5 суток (диапазон = 115-380 суток). Физическая характеристика членов основных экипажей и продолжительность космических миссий представлена в таблице.
Индивидуальные характеристики участников основных экспедиций и длительность космических полетов на Орбитальном научно-исследовательском комплексе «МИР»
|
Возраст, лет |
Рост, см |
Масса, кг |
Длительность полета, сутки |
|
54 |
167 |
79.3 |
115 сут. 08 час. 43 мин |
|
43 |
182 |
81.0 |
196 сут. 17 час. 26 мин. |
|
40 |
175 |
83.0 |
196 сут. 17 час. 26 мин. |
|
47 |
170 |
77.0 |
207 сут. 12 час. 02 мин. |
|
40 |
182 |
83.0 |
198 сут. 16 час. 31 мин. |
|
42 |
182 |
70.0 |
188 сут. 20 час 16 мин. |
|
50 |
174 |
86.0 |
379 сут. 14 час. 51 мин. |
До полета все члены экипажей были подробно информированы о целях и методах исследования сократительной функции мышц-разгибателей стопы, ознакомлены с процедурами исследования произвольных и электрически вызванных сокращений мышцы, о риске и важности настоящего исследования и после этого каждый космонавт подписал информированное согласие на участие в эксперименте. Все экспериментальные процедуры были выполнены в соответствии с Хельсинской Декларацией 2004 г. по правам человека на участие в эксперименте в качестве испытуемого и программа исследований была одобрена комиссией по биомедицинской этике при Институте медико-биологических проблем РАН и Центре подготовки космонавтов имени Ю.А. Гагарина.
Тестирующая процедура и измерения
Тендометрический динамометр. Экспериментальная установка для регистрации электрических и произвольных сокращений отдельной мышцы у человека in situ была описана ранее [6]. Коротко, обследуемый удобно располагался на специальном стуле при стандартной позиции – угол в тазобедренном, коленном и голеностопном суставах составлял ~ 90 °. Жесткая фиксация суставов конечности обеспечила изометрический режим сокращения мышцы. Позиция стула и фиксирующих устройств суставов конечности подбирались индивидуально для каждого космонавта и затем фиксировались, для воспроизведения этих параметров после возвращения космонавта из длительной космической миссии. Все тестирующие процедуры были выполнены на «ведущей» конечности космонавта.
Динамометр, представляющий стальное кольцо с вмонтированными в него тензодатчиками, плотно прижимался к Ахиллову сухожилию мышцы. Механическая деформация динамометра при сокращении мышцы преобразовывалась в электрический сигнал, и после усиления усилителем (тип «АНЧ-7м»; СССР) регистрировали на светолучевом осциллографе (тип «К-115», СССР). Степень давления датчика была постоянной для всех космонавтов и составила 5 кг. Динамометр до и после исследования калибровался, нагружая его внешними грузами разной массы.
Сократительные свойства ТМГ измерялись дважды – за 30 суток до старта и на 3-5-сутки после возвращения космонавтов из космической экспедиции на Землю. Протокол испытаний сократительных функций ТМГ до и после полета был идентичным.
Стимуляция
Для стимуляции n. tibialis использовали универсальный нейро-мышечный стимулятор (тип «ЭСУ-1», СССР) с изолирующей приставкой. Использовали монополярный электрод – активный электрод (катод, стальной шарик Ø 1 см) устанавливали в подколенной ямке (место наименьшего сопротивления), а пассивный (анод, Ag/AgCl пластина 6 х 4 см) – на нижней трети передней поверхности бедра. Положение стимулирующих электродов подбирали так, чтобы при некоторой «минимальной» силе раздражения регистрировать (по электромиографическому залпу m. soleus) наибольший прямой ответ мышцы (М-ответ). В дальнейшем силу раздражения увеличивали в 1,5-2 раза, что позволяло использовать супрамаксимальную силу раздражения (сила на 30-40 % больше той «минимальной» силы, при которой впервые достигается максимальный Мответ).
Регистрация электромиограммы
Для регистрации электромиограммы использовали поверхностные биполярные Ag/AgCl электроды (Ø 8 мм и с междуэлектродным расстоянием 25 мм), которые были помещены по средней линии брюшка камбаловидной мышцы «ведущей» конечности на уровне ниже 2 см головок икроножных мышц. Земляной электрод (Ag/AgCl пластина размеров 7.5 х 6.5 см) был помещен в проксимальной части голени между отводящими и раздражающим электродами. Для лучшего электрического контакта с кожей биполярные Ag/AgCl электроды были заполнены электродным гелем. Дополнительно, для уменьшения межэлектродного импеданса до 5 kW поверхность кожи в месте установки Ag/AgCl электродов была тщательно выбрита, обработана абразивной пастой и очищена раствором спирта с эфиром в пропорции 1 : 4 [1]. Для усиления электромиографического сигнала использовали усилитель (тип УБП-1-02, СССР) с выносным катодным повторителем. Усиленный сигнал визуально контролировали на экране запоминающего осциллоскопа (тип С8-9А, СССР) и синхронно регистрировали на светолучевом осциллографе (тип К-115, СССР).
Силовые свойства
Механические параметры произвольных и электрически вызванных сокращений ТМГ оценивали методом тендометрии [9] с использованием тендометрического динамометра [6]. Изометрические одиночные и тетанические сокращения ТМГ вызывали электрическим раздражением n. tiblalls, используя прямоугольные импульсы длительностью 1 мс супрамаксимальной силы от универсального нейро-мышечного электростимулятора (тип «ЭСУ-1», СССР) через изолирующую приставку. Применяли одиночное раздражение и тетаническую стимуляцию n. tiblalls частотой 150 имп/с [6].
Перед выполнением произвольного изометрического сокращения ТМГ каждого космонавта инструктировали, как реагировать на звуковой сигнал «сократить максимально сильно». При этих условиях космонавту сообщалась величина развиваемой произвольной силы (рис. 1, Б), а также разрешался зрительный контроль за величиной развиваемого усилия по стрелке динамометра. Каждый космонавт выполнял от 3 до 4 попыток разделенных периодом отдыха не менее 1 мин и наибольшая величина из трех попыток принималась за показатель максимальной произвольной силы (МПС).
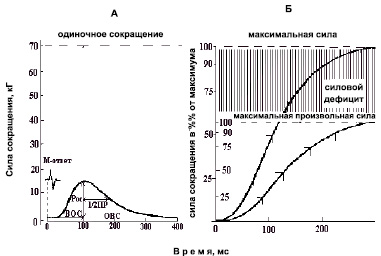
Рис. 1. Схема параметров мышечного сокращения. Пример тендограммы развития изометрического одиночного сокращения (А), электрически вызванного тетанического и произвольного сокращения (Б) мышцы с последующей схемой расчета параметров механических ответов мышечного сокращения: ВОС – время одиночного сокращения, 1/2 ПР – время полурасслабления, ОВС – общее время сокращения, Рос – сила одиночного сокращения. По оси абсцисс: время, мс; по оси ординат – сила сокращения – абсолютная, кг (левая шкала) и относительная, % от максимума (правая шкала)
По тендограмме изометрического одиночного сокращения мышцы (рис. 1, А) , в ответ на одиночный электрический импульс, нанесенный на n. tibialis, измеряли силу (пик амплитуды) одиночного сокращения (Рос) ТМГ. Максимальная сила (Ро) сокращения ТМГ (рис. 1, Б) определялась по тендограмме вызванного сокращения в ответ на электрическое тетаническое (частота 150 имп/с) раздражение n. tibialis [25, 33]. Общая длительность электрически вызванного сокращения мышцы составляла не более ~ 0,5 с.
Для количественной оценки степени совершенства центрально-нервных координационных механизмов управления произвольным (мышечным) движением рассчитывали величину силового дефицита (Рд), определяемую по разнице между Ро и МПС, выраженной в процентах от величины Ро [6] и, чем меньше величина Рд, тем выше степень произвольного использования силовых возможностей мышечного аппарата [12].
Для оценки динамики изменения «внутренней силы» контрактильных элементов мышцы (36) использовали метод парного раздражения [11], когда второй супрамаксимальный импульс наносился на n. tibialis через 3, 4, 5, 10 и 20 мс после первого и прирост напряжения в ответ на второй стимул выражался в процентах к Рос [6, 10].
Скоростные свойства
По тендограмме изометрического Рос ТМГ (рис. 1, А) в ответ на одиночный электрический импульс, приложенный к n. tibialis, рассчитывали время от момента нанесения электрического стимула (артефакт раздражения) до пика Рос (время одиночного сокращения – ВОС); время от пика Рос до половины расслабления (время 1/2 ПР) сокращения и время от момента нанесения электрического импульса до полного расслабления (общее время сокращения – ОВС). Точность измерения составляла – 2 мс.
Величину тетанического индекса (ТИ) определяли отношением Ро / Рос.
Скоростно-силовые свойства
Каждого космонавта тщательно инструктировали, как реагировать на звуковой сигнал – «сократить мышцу максимально быстро и сильно». Регистрируемое произвольное сокращение ТМГ принималось как «взрывное» баллистическое сокращение. Скорость нарастания, или иначе градиент, изометрического напряжения ТМГ рассчитывали по тендограмме изометрического произвольного сокращения, используя «относительные» показатели, т.е. время достижения 25, 50, 75 и 90 % уровня от максимума [22, 38, 34]. Аналогично по тендограмме электрически вызванного сокращения при стимуляции n. tibialis с частотой 150 имп/с [6] определяли время нарастания электрически вызванного сокращения (рис. 1, А). Точность измерения составляла – 2 мс.
Максимальную скорость развития произвольного (dPпс/dt) и электрически вызванного сокращения (dPвс/dt) определяли путем дифференцирования механических (аналоговых) ответов мышцы.
Статистика
При обработке полученных результатов исследования использовали общепринятые статистические методы: рассчитывали среднюю и стандартную ошибку средней (М ± m). Кроме того, различие между фоновыми (контрольными) показателями и показателями, зарегистрированными после завершения космической миссии, оценивали с помощью параметрического t–критерия Стьюдента и величину значения р < 0.05 принимали как существенную.
Результаты исследования и их обсуждение
Космический полет и силовые свойства. Средние данные изменения силовых свойств ТМГ после продолжительного пребывания в условиях невесомости графически представлены на рис. 2, A. Анализ полученных результатов обнаружил изменения силовых характеристик мышцы. Так, величина изометрической Рос после космического полета несущественно увеличилась в среднем на 14.8 % (до 10,8 ± 0,8 кг после 12,4 ± 1,8 кг; р > 0,05) и существенно уменьшились МПС в среднем на 41.7 % (до 53,0 ± 6,8 кг после 30,9 ± 5,1 кг; р < 0.01) и Ро в среднем на 25.6 % (до 75,3 ± 6,7 кг после 56,0 ± 3,8 кг; р < 0.05).
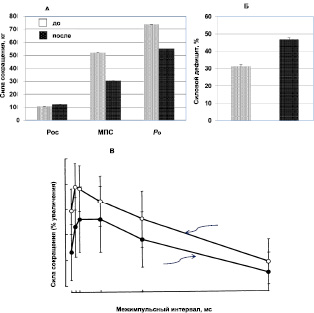
Рис. 2. Влияние продолжительных космических полетов на силу сокращения мышцы: А. Максимальный ответ одиночного сокращения (Pос), максимальная произвольная сила (МПС), максимальное электрически вызванное тетаническое сокращение (Ро; частота 150 имп/с). Б. Силовой дефицит (Рд), до и после космических полетов (средняя группа). В. Сила сокращения трехглавой мышцы голени при парном раздражении с разными интервалами между импульсами
Величина МПС ТМГ как до, так и после космического полета была постоянно меньше, чем Ро этой же мышцы и величина Рд, указывающая на степень совершенства центрального звена управления мышечным аппаратом, после полета увеличилась в среднем на 49.7 % (до 32.2 ± 4.6 после 46.7 ± 5.4 %; р < 0.001) (рис. 2, Б).
При анализе индивидуальных показателей величины МПС обнаружено, что силовые сократительные свойства ТМГ, в целом, имеют однонаправленные изменения – снижение силовых свойств (рис. 3). Снижение МПС было обнаружено у каждого космонавта по сравнению с фоновыми данными, которые были значительно выше перед полетом по сравнению с послеполетными показателями (р < 0.05), но степень снижения разнилась. Так, у 3 космонавтов (№ 3, 5 и 6) было обнаружено снижение МПС в пределах от 20 до 30 %, а у 2 космонавтов (№ 4 и 6) – более 60 %.
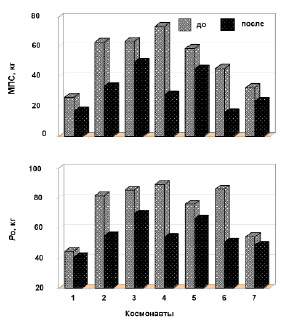
Рис. 3. Изменение максимальной произвольной силы (МПС) и максимальной электрически вызванной тетанической силы сокращения (Ро) мышцы. Гистограммы представляют индивидуальные величины МПС и Ро космонавтов до и после
Анализ изменения собственно-силовых свойств ТМГ показал снижение величины Ро у всех космонавтов (рис. 3). Уменьшение Ро было обнаружено для каждого космонавта в пределах от 8 до 41 % и лишь у 2 космонавтов (№ 1 и 7) было относительно меньшее снижение Ро (от 13 до 18 %, соответственно). Для всех космонавтов Ро была постоянно и значительно выше в предполетных условиях, чем в послеполетном состоянии.
Анализ данных не обнаружил связи между исходной величиной МПС сокращения ТМГ и степенью снижения силовых свойств мышцы после космического полета. Так, например, если космонавты № 2, 3 и 4, имеющие до космического полета самые высокие и в среднем примерно одинаковые показатели МПС, то после полета обнаруживают различия в степени снижения силы сокращения мышцы (например, космонавты № 2 и 3). Аналогично не обнаружено таких связей и для Po (рис. 3; космонавты № 2, 3, 4 и 6).
Средние данные изменения силы сокращения ТМГ при парном раздражении с разными межимпульсными интервалами графически представлены на рис. 2, C. Анализ результатов показал, что наибольшая сила сокращения при этих условиях отмечается при интервале между интервалами 4–10 мс и уменьшение или увеличение межимпульсного интервала сопровождались значительным снижением силы сокращения. После продолжительного космического полета относительный прирост силы сокращения на второй импульс был значительно ниже по сравнению с исходной величиной (р < 0.01-0.001).
Космический полет и скоростные свойства. Данные изменений среднего времени развития изометрической Рос ТМГ, как обратной величины скорости сокращения, после космического полета представлены графически на рисунке 4, A. Как следует из анализа полученных данных отмечается «замедление» скорости сокращения мышцы и увеличение времени 1/2 ПР. Так, после космического полета ВОС увеличилось в среднем на 7.7 % (130 ± 2 против 140 ± 6 мс; р < 0.05), а время 1/2 ПР – уменьшилось в среднем на 20,6 % (97 ± 4 против 77 ± 4 мс; р < 0.05). ОВС увеличилось после космического полета в среднем на 7.5 % (456 ± 25 против 490 ± 31 мс; р < 0.05).
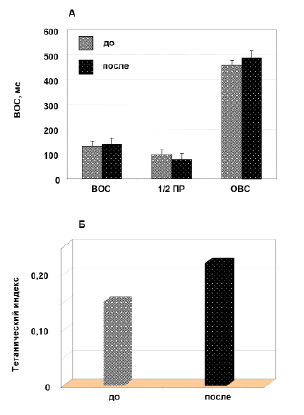
Рис. 4. Влияние продолжительных космических полетов на скоростные свойства мышцы: A. Время одиночного сокращения (ВОС), время полурасслабления (1/2 ПР) до и после полета. B. Тетанический индекс
Другим косвенным показателем, характеризующим скоростные свойства мышц, является величина ТИ, т.е. отношение Рос/Ро (рис. 4, Б). Анализ представленных данных показывает значительное увеличение этого показателя в среднем на 46.7 % (до 0.15 ± 0.01 после 0.22 ± 0.02).
Как следует из анализа индивидуальных данных, ВОС ТМГ под влиянием продолжительной космической миссии увеличивается, но вместе с тем следует отметить, что скорость сокращения мышцы изменяется неоднозначно (рис. 5). Так, увеличение ВОС было обнаружено для 5 космонавтов в пределах от 8 до 24 %, тогда как 2 космонавта (№ 3 и 4) показали незначительное уменьшение на 6 и 12 %, соответственно.
Сравнительный анализ изменения индивидуальных данных времени 1/2 ПР ТМГ у космонавтов до и после полета также показал неоднозначные изменения. Так, 7 космонавтов показали уменьшение времени 1/2 ПР в пределах от 2 до 37 %, тогда как космонавт № 5 обнаружил незначительное (2 %) изменение (рис. 5).
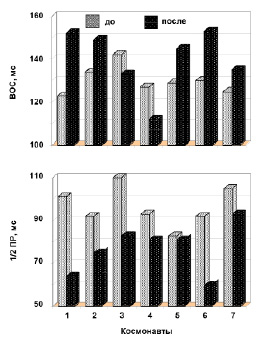
Рис. 5. Изменение скоростных свойств мышцы. Гистограммы представляют индивидуальное величины ВОС и 1/2 ПР космонавтов до и после полета
Космический полет и скоростно-силовые свойства. Снижение МПС после продолжительного космического полета сопровождалось существенным замедлением времени (обратная величина скорости) развития произвольного изометрического сокращения мышцы. Данные об изменении времени нарастания изометрического сокращения ТМГ после космического полета графически представлены на рис. 6. Уменьшение МПС (в среднем на 42 %) было связано с существенным замедлением скорости, или градиенте развития изометрического произвольного сокращения мышцы (рис. 6, левая панель р < 0.01-0.001); максимальная dPпс/dt была существенно снижена (в среднем на 36.2 %), когда была выражена в относительных единицах, т.е. как процент от МПС (рис. 6, вставка; р < 0.001).
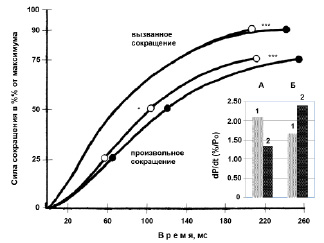
Рис. 6. Влияние продолжительных космических полетов на скоростно-силовые свойства мышцы (левая панель) и максимальную скорость развития сокращения (вставка; А – произвольное сокращение; Б – вызванное сокращение: 1 – до; 2 – после)
Анализ кривой сила–время электрически вызванных сокращений не обнаружил существенных различий (рис. 6, левая панель; р > 0.05), в то время как максимальная dPвc/dt была существенно увеличена (в среднем на 43.7 %) после космических полетов (рис. 6, вставка; р < 0.001). Эта механическая диссоциация указывает, что были изменены некоторые вне мембранные процессы.
Сравнение космического полета с постельным режимом. В предыдущей работе мы исследовали сократительные ответы ТМГ у испытуемых после пребывания в течение 120 суток на жестком постельном режиме (6, 36). Сравнение данных, полученных в настоящей работе с данными исследований предыдущей работы, графически представлены на рисунке 7. Степень изменения сократительных свойств ТМГ, по-видимому, одинакова в этих двух моделях. Данные указывают, что нет никаких относительных различий между этими двумя моделями для МПС, Pо, Pд, ВОС и 1/2 ПР. С другой стороны, наши данные демонстрируют, что физическая тренировка, используемая во время космических полетов, не была абсолютно успешной в предотвращении изменений в нервно-мышечной работоспособности, что указывает на необходимость разработки более эффективных подходов в тренировочном процессе.
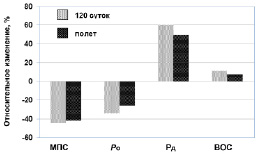
Рис. 7. Влияние продолжительного космического полета и 120-суточного постельного режима на сократительные свойства трехглавой мышцы голени. Данные постельного режима получены от (36)
Целью настоящего исследования было определить влияние продолжительного пребывания в условиях микрогравитации (космический полет) на произвольные и непроизвольные (электрически вызванные) сокращения ТМГ у здоровых мужчин-космонавтов. Основным результатом настоящего исследования было снижение сократительных свойств ТМГ после продолжительного космического полета. Изменение в функциях мышцы под воздействием экспериментальных условий может быть обусловлено либо изменениями в сократительных процессах, либо в нервной (моторной) команде.
Использование электрически вызванных одиночных и тетанических сокращений хорошо обосновано для исследований сократительных функций мышц у человека, поскольку параметры электрически вызванного сокращения мышцы – сила, скорость и длительность сокращения (при выбранном способе раздражения) – обусловлены функциональными свойствами самой исследуемой мышцы.
Результаты настоящего исследования демонстрируют, что сократительные свойства ТМГ изменяются во время продолжительного космического полета на борту станции «МИР». Эксперименты, использующие различные модели с разгрузкой мышц или реальный полет, показали изменения в генерации изометрической силы сокращения мышцы, скорости и связи сила-скорость [2, 4, 17]. По-видимому, эти изменения являются результатом отсутствия нормальной весо-нагрузочной активности постуральных мышц нижних конечностей, вызванной сниженными условиями силы гравитации на борту космической станции.
Влияние космического полета на силовые свойства
Мы ожидали, что изменения в изометрической Pос будут соответствовать изменениям, наблюдаемым с МПС, в частности уменьшением. Напротив, мы наблюдали несущественное увеличение Pос после продолжительного космического полета, что согласуется с ранее полученными данными [6, 23]. Эти изменения, по-видимому, можно рассматривать, как ответную реакцию мышечного аппарата на изменение жесткости (растяжимости) мышцы. Уменьшение растяжимости (жесткости) мышцы, которая отмечалось после неупотребления/космического полета [7, 42] должно вносить измения в характеристики развиваемой изометрической Рос. Напротив, после космического полета величина, характеризующая собственно-силовые свойства ТМГ (Po), уменьшилась и, таким образом, увеличение величины ТИ после космического полета отражает изменения в растяжимости (жесткости) мышцы. Мы допускаем, что в нашем эксперименте космический полет явился причиной увеличения растяжимости мышцы.
После продолжительного космического полета электрически вызванное сокращение (Ро) существенно уменьшилось (на 26 %). Показатель Ро есть прямая мера, отражающая способности мышцы генерировать силу, и отражает число активных связей между актиновыми и миозиновыми нитями [14]. Неупотребление, как показано, продуцирует снижение Ро [6, 22]. Это снижение отражает уменьшение числа активных поперечных мостиков и, как следствие снижение работоспособности. Можно предположить две гипотезы снижения Ро мышцы. Во-первых, нельзя исключить, что общее число поперечных мостов мышцы было снижено после космического полета. Во-вторых, продукция силы, развиваемая каждым поперечным мостиком была также уменьшена. Однако, когда развиваемая сила была выражена на площадь поперечного сечения, то сила оставалась неизменной после неупотребления [54]. Это означает, что второе предположение должно быть отвергнуто. Таким образом, уменьшение Ро объясняется уменьшением максимального числа поперечных мостов в ТМГ.
В целом механизмы, ответственные за снижение силы сокращения мышцы при воздействии микрогравитацией, остаются не вполне ясными. Снижение силы, возможно, вызвано изменениями в архитектуре мышцы, т.е. длине мышцы, длине и угле наклона мышечных волокон [34, 58]. Следовательно, связь угол сустава, длина мышцы и угол наклона волокон являются высокоспецифичными для мышцы. Архитектура мышцы может влиять на функциональные характеристики мышцы, например, максимальную скорость сокращения и величину Ро [48]. Кроме того, принимая во внимание, что ТМГ в условиях невесомости не растянута и не напряжена, то создаются условия для физиологического укорочения и можно предположить, что в этом случае общее число последовательно-соединенных саркомеров было уменьшено [25]. Последнее, возможно, способствовало снижению толщины мышцы и угла наклона волокон. Действительно во время космического полета космонавты принимают «флексорное» положение [21].
Более того, МПС мышцы зависит от таких факторов, как мотивация космонавтов в задаче «развить максимальное усилие». Величина МПС, полученная до полета, соответствует литературным данным [15, 31, 39, 54] и как показано другими исследователями, которые также принимали участие в исследованиях на космической станции «МИР» [16, 60], наши после полетные данные указывают, что МПС была снижена у каждого космонавта. Было обнаружено, используя технику ЯМР, уменьшение массы мышц нижних конечностей в среднем на ~ 11 %, что объясняет уменьшение площади поперечного сечения мышцы [60]. Наши после-полетные индивидуальные данные космонавта указывают на существенное уменьшение МПС. Большее снижение МПС можно предположить определяется изменениями характера рекрутирования двигательных единиц (ДЕ) и изменениями порядка рекрутирования ДЕ [5, 45]. Подтверждением этого является снижение после космического полета мощности ЭМГ сигнала [49] и указывает, что микрогравитация оказывает существенное воздействие в нормальный нервный паттерн.
Намного большее снижение МПС (в среднем на 42 %) по сравнению с изменением в Ро (в среднем на 24 %) после продолжительного космического полета может указать на неспособность центральной нервной системы нормально активировать ТМГ. Было ли это из-за нехватки мотивации со стороны космонавтов или в непроизвольном уменьшении нервного драйва трудно различить. Дополнительно, снижение в максимальной мощности после продолжительного (180 суток) космического полета было объяснено уменьшением в нервной посылке [16]. Принимая во внимание, что наши космонавты были высоко мотивированы и не имели никакого дискомфорта во время развития МПС, то предположение о низкой мотивации, как причине развития низкой величины МПС, должно быть отвергнутым. Увеличение Рд подтверждает снижение центрального драйва контроля произвольного управления мышцей нервной системы. Действительно, как отмечалось, непосредственно во время бездеятельности электромиографическая (ЭМГ) активность мышцы была значительно снижена во время развития МПС [13, 22, 37, 51]. Кроме того, анализ изменения амплитуды ЭМГ мышцы после бездеятельности позволяет предполагать, что меньшее число ДЕ в мышце было активировано при неупотреблении [51] и максимальная частота импульсации ДЕ была уменьшена [22]. Фактически, уменьшение максимальной частоты импульсации подразумевает рекрутирование большего числа ДЕ, чтобы развить ту же самую величину силы сокращения мышцы в послеполетных условиях.
С другой стороны, уменьшение максимальной частоты импульсации можно объяснить изменениями проприоцептивного входа на мотонейроны в связи с разгрузкой мышц [40, 44]. Изменения в моторном контроле произвольных движений подтверждается существенным уменьшением ЭМГ активности мышцы (снижением ЭМГ мощности по сравнению с потерей в массе мышцы) при выполнении «взрывной» силы [15, 16]. Последнее позволяет предположить, что микрогравитация вызывает реорганизацию в порядке рекрутирования ДЕ [49]. Эти изменения в моторном контроле у космонавтов/астронавтов после космического полета могут быть одним из лимитирующих факторов, определяющих величину снижения максимального суставного момента, развиваемого мышцами-разгибателями, и максимальной мощности [2, 16]. Дополнительными факторами в снижении силы мышцы является изменение архитектуры мышцы и мышечно-сухожильного комплекса, как ранее было показано после полета астронавтов [42] и после постельного режима [34, 41].
Мы изучили сократительные свойства ТМГ, полученные у взрослых испытуемых до и после 120 суток постельного режима [6, 36]. Во время постельного режима протокол исследований сократительных свойств ТМГ испытуемых был аналогичным пред- и пост-полетным обследованиям членов космических миссий. Различие в величине снижения МПС и Ро ТМГ во время 120-суточного постельного режима и космического полета было не больше, чем на несколько процентов. Таким образом, результаты, в целом, указывают, что механизмы, ответственные за генерацию силы сокращения ТМГ, сходны как при постельном режиме, так и при космическом полете. Однако необходимо отметить, что одно различие между исследованиями при постельном режиме и после космического полета было в том, что космонавты во время полета выполняли упражнения преимущественно аэробной направленности (тредмил, велоэргометр). Поэтому трудно сопоставить данные между постельным режимом и космическим полетом. Мы согласились бы с заключением, если бы испытуемые не выполняли физическую тренировку во время постельного режима. Во время постельного режима испытуемые действительно выполняли физическую тренировку близкой к программе, предложенной в полете. В результате этого величина МПС (особенно) и Ро незначительно снизились (всего лишь на 3 и 9 %, соответственно), и значительно уменьшился Рд (на 10 %). Полученные результаты показали, что в ответ на физическую тренировку в условиях постельного режима происходит адаптация как нервная, так и мышечная, но больше нервная [51].
Влияние космического полета на скоростные свойства. Результаты показали, что микрогравитация оказывает воздействие на медленно сокращающуюся группу мышц [6, 8, 10]. Данные дополняют ранее полученные результаты влияния как кратковременной [6, 10], так и продолжительной разгрузки на механические характеристики ТМГ [6, 40]. Функциональными компонентами длительности Рос являются ВОС и время 1/2 ПР. Существенное увеличение ВОС и снижение времени 1/2 ПР были обнаружены во время 120-суточного постельного режима [6]. В настоящем исследовании ВОС ТМГ до и после полета составило ~ 8 %. Основным механизмом изменения ВОС и времени 1/2 ПР может быть конкурирующие между собой взаимодействия факторов, связанных с активацией (кинетика освобождения Ca2+) цикла образования поперечных мостиков, потреблением Ca2+ саркоплазматическим ретикулумом (СР) и изменением активной фракции (мышечные волокна) или пассивной фракции (сухожилия) серии эластического компонента.
Возможным объяснением изменения ВОС под воздействием реальной микрогравитации может быть относительно большая степень атрофии волокон типа I, которые составляют большинство в ТМГ [33]. Однако, как известно микрогравитация продуцирует атрофию как быстрых, так и медленных мышц, вызывая специфические изменения в сократительных свойствах волокон [27]. Дополнительно, можно допустить, что уменьшение времени 1/2 ПР ТМГ происходит, вероятно, благодаря увеличению числа СР Са2+-АТФ-азы помп, как наблюдается в предыдущих исследованиях при разгрузке мышц конечностей [52].
Дополнительным фактором является распределение типа волокон. Показано, что после космического полета процент волокон несущественно изменяется. Отмечалось после космического полета небольшое увеличение процента быстросокращающихся волокон типа IIa [24, 57]. В этой связи можно было бы ожидать увеличение скорости развития вызванного одиночного сокращения мышцы. Однако, как показывают наши данные, ВОС мышцы после космического полета обнаруживает тенденцию к увеличению. С этими данными согласуются ранее полученные данные в условиях, моделирующих невесомость [6, 37].
Быстрая природа изометрических изменений, т.е. длительность Рос, может быть связана с изменениями в функции СР [18, 26]. Первичным фактором (механизмом) объяснения этих изменений может быть сокращение уровня, при котором Ca2+ отделяется от миофибриллярных белков [18]. Диссоциация происходила бы более медленно, если бы скорость потребления Ca2+ СР была снижена. Такое уменьшение было обнаружено при неупотреблении [35]. Снижение скорости отделения Ca2+ от миофибриллярных белков, как можно было ожидать, не только увеличит время кривой ответа одиночного сокращения, но также позволит генерировать большую величину силы, т.к. продолжатся формироваться поперечные мостики, поскольку Ca2+ находится в саркоплазме. Эти эффекты на СР было бы трудно наблюдать, поскольку эффекты на Pос будут замаскированы атрофией, но представляют интерес при условии, что изменения одиночного сокращения происходят благодаря изменениям в СР. Изменение кинетики механических ответов при парном раздражении (см. рис. 2, C) может быть также объяснением изменения развития Ca2+ кинетики в мышце. При любом межимпульсным интервале относительное увеличение силы сокращения после продолжительного космического полета было значительно меньше по сравнению с предполетной величиной.
Влияние космического полета на скоростно-силовые свойства. Скорость развития вызванного сокращения (форма кривой сила – время) в ответ на электрическое раздражение нерва с частотой 150 имп/с, используя «относительные» показатели, незначительно изменяется после космического полета, что согласуется с наблюдениями [59] об относительно постоянной механике развития тетанического сокращения и современной теории (поперечных мостиков) мышечного сокращения [53]. Как ранее было показано, в условиях моделирующих продолжительный космический полет (постельный режим в течение 120 суток), скорость развития электрически вызванного тетанического сокращения ТМГ существенно не изменяется [6] и это наблюдение хорошо согласуется с данными, полученными ранее [59]. Исходя из посылки, что форма кривой сила – время определяется скоростью формирования и разрыва связи поперечных мостиков [53], которая пропорциональна активности мио–АТФ-азы [54], то можно предположить, что цикличность поперечных мостиков и активность мио–АТФ-азы мало изменяется (или совсем не изменяется) под воздействием микрогравитации. Поэтому можно предположить, что пассивная активность мио–АТФ-азы причина изменения кинетики сокращения после невесомости. Это поддерживает гипотезу, что причиной изменения изометрических сократительных свойств мышц, находящихся в условиях микрогравитации, является не изменения сократительных белков, а возможно определяет любой другой фактор/ы, возможно, от изменения в функции СР. Увеличение нормализованной dPвс/dt, которая была зарегистрирована после продолжительного космического полета (см. рис. 6), согласуется с данными, что активность мио–АТФ-азы и максимальная скорость укорочения были увеличены в результате космического полета, что согласуется с наблюдениями [56, 57]. Поэтому можно заключить, что неупотребление мышц (например, невесомость или постельный режим) в продолжительном космическом полете имеет небольшое влияние на цикличность поперечных мостиков или на активность миозина [57].
В дополнение, как известно, время кривой сила – время электрически вызванного тетанического сокращения действительно не изменялось, тогда как максимальная скорость dPвс/dt была увеличенной. Причина, увеличенной скорости укорочения разгруженной мышцы, вызванная космическим полетом, неизвестна. Однако увеличение скорости укорочения, как результат микрогравитации, может быть следствием геометрических изменений в структурах мышцы, т.е., увеличение пространства миофиламентов [57] или в уменьшение угла наклона волокон мышцы [34, 58].
С другой стороны, увеличение максимальной dPвс/dt укорочения ТМГ после полета могло произойти, по крайней мере, по трем возможным механизмам: i) увеличение быстрого типа тяжелых цепей миозина (MHC); ii) de novo более быстрых изоформ MHC, и/или iii) увеличение быстрых цепей легкого миозина (MLC). Действительно была обнаружена хорошая корреляция между составом изоформ MHC и максимальной скоростью укорочения разгруженной целой мышцы (26) или отдельного волокна [43, 56]. Следовательно, 44 % увеличение максимальной dPвс/dt ТМГ могло произойти из-за соответствующего увеличения содержания относительно быстрого типа изоформ MHC. Также возможно, что частичное увеличение в максимальной dPвс/dt укорочения, возможно, произошло из-за изменений в существенном составе изоформ MLC [57] и возможно, из-за потери актиновых нитей [50]. Эти структурные изменения могут быть ответственными за послеполетное увеличение максимальной dPвс/dt сокращения и уменьшение жесткости мышцы [57], что имеет большее влияние на скорость укорочения, чем на силу [46, 57].
В заключение, настоящие результаты показывают, что механические свойства ТМГ человека были изменены после продолжительного космического полета. Сравнение механических изменений, зарегистрированных во время произвольных и электрически вызванных сокращений (для всех семи космонавтов после полета МПС продуцирует большее снижение, чем Ро), предполагает, что невесомость не только изменяет периферические процессы, связанные с сокращениями, но также и изменяет центральную и/или нервную команду сокращения.
В добавлении, снижение сократительных функций мышц, полученных от членов экипажей космических миссий, поддерживает представление, что защита скелетных мышц во время продолжительного космического полета требует физической подготовки. Кроме того, предполагается, что физическая подготовка во время космического полета была выполнена с умеренной интенсивностью, но большого объема и, таким образом, улучшая аэробную производительность, а не мощность сократительных свойств. Высокоинтенсивные и/или «взрывные» упражнения, можно предположить, являются многообещающими в предотвращении атрофии мышц и «слабости». Кроме того, анализ тренировочного процесса, используемого в космическом полете, не обнаружил специальных упражнений, направленных на тренировку мышц-разгибателей стопы, что может быть дополнительным фактором в снижении сократительных функций мышцы.
Автор выражает благодарность всем членам экипажа космической станции «МИР», которые любезно согласились на участие в этих исследованиях и без их усилий этот проект был бы невыполним, а также благодарит медицинский и инженерный штат ЦПК имени Ю.А. Гагарина (Звездный городок, Московский регион) за их вклад в организацию исследования, а также многочисленным неназванным сотрудникам, которые помогали в осуществлении нашего исследования.
Автор также выражает особую благодарность И.Б. Козловской за поддержку данного научного направления при проведении исследований в рамках совместных российско-американских программ «MIR-SHUTTLE» и «MIR-NASA».
Данное исследование было поддержано ГНЦ – Институтом медико-биологических проблем РАН.
Библиографическая ссылка
Коряк Ю.А. СОКРАТИТЕЛЬНЫЕ СВОЙСТВА ТРЕХГЛАВОЙ МЫШЦЫ ГОЛЕНИ У ЧЕЛОВЕКА ПОСЛЕ ПРОДОЛЖИТЕЛЬНОГО ПРЕБЫВАНИЯ В УСЛОВИЯХ НЕВЕСОМОСТИ НА БОРТУ ОРБИТАЛЬНОЙ СТАНЦИИ «МИР» // Международный журнал прикладных и фундаментальных исследований. – 2013. – № 8-3. – С. 27-41;URL: https://applied-research.ru/ru/article/view?id=3868 (дата обращения: 19.04.2024).