

Необходимость постоянного увеличения производства продуктов питания и кормов для животных повысила потребность в расширении производства сои. Основные продукты из сои – это соевая мука и соевое масло, которые используются для приготовления кондитерских изделий, наполнителей, заменителей мяса, молока, сыра, диабетических продуктов, майонеза, маргарина. Создание продуктивных сортов сои может не только обеспечить людей продуктами питания, а животноводство – полезными кормовыми продуктами, но и помочь свести к минимуму импорт кормовых ресурсов. Но, несмотря на создание перспективных сортов и повышение урожайности, потери, вызванные абиотическими и биотическими стрессовыми факторами, в частности засухой, также занимают немало места [1]. В селекции устойчивых сортов и форм важна разработка ранней диагностики засухоустойчивости растений. Несмотря на большое потребление воды соя растет и при определенном дефиците влаги. Потребность сои в воде по фазам развития неодинаковая [2, с. 148]. В растительном организме засуха способна оказывать сильное влияние на ход физиологических процессов, но в основе лежит влияние засухи на водный режим. Изучение параметров водного режима составляет основу определения засухоустойчивости растений.
В связи с изложенным цель наших исследований заключалась в оценке параметров водного режима и содержания фотосинтетических пигментов у образцов сои в условиях засухи.
Материалы и методы исследования
Использованные в исследовании семена 21 сорта сои (Glycine max L.): Чукурова-3, Чукурова-4, Чукурова-5, Чукурова-6, Чукурова-7, Чукурова-8, Чукурова-9, Чукурова-12, Чукурова-13, Чукурова-14, Канада-1, Канада-4, Канада-5, Канада-6, Канада-7, Уманская, Антониа, Ангелика, Аяз, Алекса, Киото – были предоставлены Национальным Генбанком Азербайджана и выращены на опытно-полевом участке института. Параметры водного режима в листьях растений определяли на фазе ветвления и цветения в условиях полива и на фазе ветвления при засухе по методике Н.Н. Кожушко [3, с. 49].
Для изучения взаимосвязи между устойчивостью генотипов сои к стрессовым факторам и содержанием хлорофилла для анализа были взяты образцы верхних листьев в фазу ветвления и в фазу цветения интактных полевых растений. Листья подвергались стрессу в лабораторных условиях в течение 24 ч. Для моделирования стресса был использован раствор 8,7 % сахарозы (7 атм), соответствующий пределу устойчивости. На спектрофотометре (UV-3100 PC) измеряли оптическую плотность хлорофилла при 665 нм (xl a) и 649 нм (xl b), каротиноидов при длине волны 450 нм [3, с. 22].
Результаты исследования и их обсуждение
Параметры водного режима сортообразцов сои определяли в условиях жаркого климата Апшерона. Определение водного дефицита выявило различие по этому показателю между образцами сои (рис. 1). У образцов сортов сои Чукурова-9, Киото, Антониа и Канада-6 наблюдался низкий водный дефицит в фазе ветвления, соответственно 10,3; 11,5; 13,8 и 14,2 %. Максимальный водный дефицит был отмечен у образца Чукурова-3 (34,2 %), высокий водный дефицит – у сортов Чукурова-4, Чукурова-7, Канада-7, соответственно 27,2; 24,7; 25,3 %.
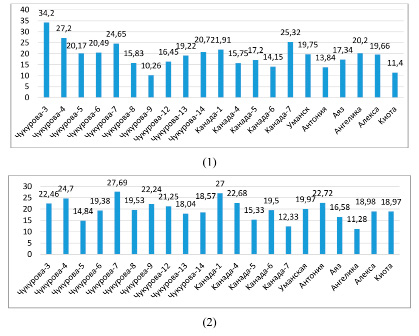
Рис. 1. Водный дефицит листьев растений сои в условиях орошения: 1 – фаза ветвления; 2 – фаза цветения (%)
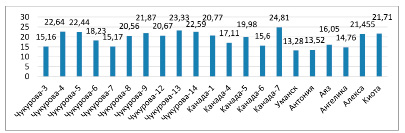
Рис. 2. Водный дефицит листьев растений сои в условиях засухи (%)
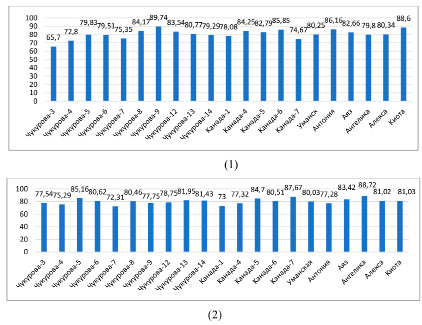
Рис. 3. Относительное содержание воды в листьях растений сои в условиях орошения: 1 – фаза ветвления, 2 – фаза цветения (%)
В фазе цветения водный дефицит был низким у образцов сортов Ангелика, Канада-7, Чукурова-5, соответственно 11,3; 12,3 и 14,8 %. Высокие показатели водного дефицита были отмечены у растений сортов Чукурова-4, Чукурова-7, Канада-1: 24,7; 27,7; 27,0 %. Во время засухи низкий водный дефицит наблюдался у сортов Ангелика, Антониа и Уманская, 14,76; 13,52 и 13,28 %, сорта Чукурова-4, Чукурова-5, Чукурова-13, Канада-7 отличились высоким водным дефицитом: 22,6; 22,4; 23,3 и 24,8 % (рис. 2).
Показатели относительного содержания воды в тканях листьев растений сортов сои варьировали в пределах 65,7 % (Чукурова-3) и 89,74 % (Чукурова-9) в фазу ветвления в условиях орошения (рис. 3). Относительная влажность в фазу цветения составила от 72,3 (Чукурова-7) до 88,7 % (Ангелика), в период засухи этот показатель находился в пределах 75,19 (Канада-7) – 86,72 % (Уманская). Относительное содержание воды выше 80,0 % наблюдалось у сортов Чукурова-5, Чукурова-6, Чукурова-8, Чукурова-13, Чукурова-14, Канада-5, Канада-6, Канада-7, Уманская, Ангелика, Аяз, Алекса, Киото в фазе цветения; Чукурова-3, Чукурова-6, Чукурова-7, Канада-4, Канада-5, Канада-6, Уманская, Антониа, Ангелика, Аяз, Алекса при засухе (рис. 4).
По водоудерживающей спoсoбности можно судить о возможности растений противостоять заcухе. У контрольных рacтений (в фазе ветвления) водоудерживающая способность колебалась в диапазоне 27,3 (Чукурова-3) – 50,6 % (Антониа), в фазе цветения – 30,21 (Чукурова-6) – 70,06 % (Канада-6), в условиях засухи – в диапазоне 33,34 (Чукурова-12) – 55,19 % (Канада-7) (рис. 5). Анализ процентного соотношения водоудерживающей способности растений, подвергнутых засухе, по сравнению с контрольными образцами, показал меньшую потерю воды у образцов сортов Канада-7 (55,19 %), Ангелика (54,27 %), Антониа (53,17 %) и Канада-1 (52,53 %) (рис. 6).
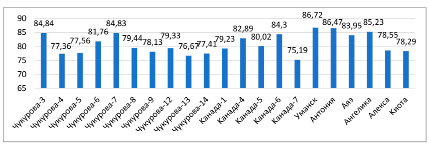
Рис. 4. Относительное содержание воды в листьях растений сои в условиях засухи (%)
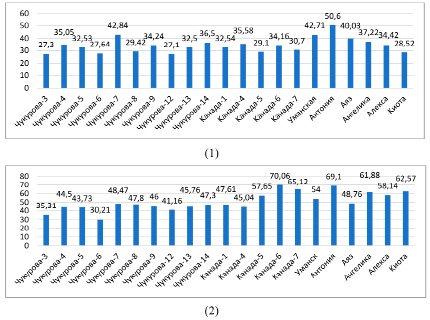
Рис. 5. Водоудерживающая способность листьев растений сои в условиях орошения: 1 – фаза ветвления, 2 – фаза цветения (%)
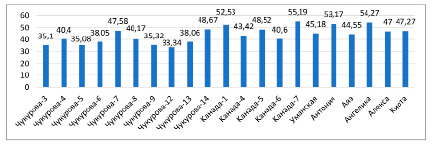
Рис. 6. Водоудерживающая способность листьев растений сои в условиях засухи (%)
При нехватке воды увеличивалась водоудерживающая способность и содержание связанной воды с осмотическими соединениями, накапливающимися в клетках. По мнению авторов, у сортов с более высокой степенью устойчивости по мере увеличения засухи усиливается способность удерживать воду в клетках растений [4]. Величина водоудерживающей способности в условиях засухи увеличилась на 4,6–6,0 % по сравнению с фазой ветвления, на 3,1–5,1 % по сравнению с фазой цветения. В фазе цветения водоудерживающая способность оказалась выше 50,0 % у 8 образцов сои: Уманская (54,0 %), Канада-5 (57,7 %), Алекса (58,1 %), Ангелика (61,9 %), Киото (62,6 %), Канада-7 (65,1 %), Антониа (69,1 %), Канада-6 (70 %). В период засухи этот показатель составил более 50,0 % у 4 образцов сортов сои: Kaнада-1 (52,53 %), Антониа (53,2 %), Ангелика (54,3 %), Канада-7 (55,2 %). В то же время по мере старения листьев увеличивается водоудерживающая способность. Это было наглядно продемонстрировано в наших экспериментах – увеличение водоудерживающей способности наблюдалось при сравнении фаз ветвления и цветения. В фазу цветения показатель водоудерживающей способности листьев увеличился на 2,9–19,5 % по сравнению с фазой ветвления.
В условиях засухи наблюдалось значительное изменение содержания фотосинтетических пигментов (таблица). За счет увеличения содержания xl a и b в условиях засухи сумма xl(a+b) составляла более 100,0 % по сравнению с контролем у образцов сортов сои Чукурова-4, Чукурова-5, Чукурова-7, Чукурова-9, Чукурова-14, Канада-1, Канада-7, Аяз, Ангелика. В вариантах опыта Чукурова-3, Чукурова-13, Канада-5, Киото, Алекса произошло снижение содержания пигментов и показатель xl(a+b) варьировал в пределах 83,1–87,99 % по сравнению с контролем. Однако, несмотря на низкое значение суммы пигментов, соотношение xl a/b превышало 100,0 %, что связано с большим уменьшением содержания хl b, чем хl а.
Исследования показали, что у большинства образцов наблюдалось увеличение содержания каротиноидов (таблица). У сортообразца Аяз оно составило 140,0 %, у сорта Канада-5 – 163,0 %, у Чукурова-14 – 175,8, у Канада-1 – 190,0 %. По-видимому, 24-часовое воздействие стрессового фактора стимулировало синтез вспомогательных пигментов. Деградация каротиноидов очень сильно проявлялась у растений сорта Ангелика (40,68 %).
Повышение и понижение соотношения хлорофилла а/b в опытных вариантах по сравнению с контролем связано с разной реакцией синтеза форм хлорофилла а и b на засуху. Фотосинтетические пигменты составляют основу фотосинтетического аппарата, поглощая световую энергию и превращая ее в химическую. Увеличение содержания хлорофилла в некоторых образцах при низких дозах стресса засухи указывает на то, что небольшой уровень стресса стимулирует синтез пигментов.
Однако по мере увеличения дозы стресса или увеличения его продолжительности в большей степени наблюдается деградация фотосинтетических пигментов. Считается, что причиной резкого снижения содержания обеих форм пигмента является нарушение активности ферментов, индуцирующих их синтез, под влиянием засухи. Важным показателем сбалансированности процессов фотосинтеза является соотношение форм хлорофилла (a/b). Так, xl a входит в состав реакционных центров фотосистем, а xl b – в состав светособирающего комплекса [5]. Следовательно, ослабление синтеза пигментных форм свидетельствует о возникновении нарушений в работе фотосистем и, как следствие, снижении эффективности процессов сбора энергии и биомассы.
Содержание фотосинтетических пигментов в листьях образцов сои при засухе
|
Образцы |
Содержание хлорофилла |
Каротиноиды |
||||||||
|
контроль |
засуха |
контр. |
засуха |
|||||||
|
a |
b |
a+b |
a/b |
a |
b |
a+b |
a/b |
|||
|
Чукурова-3 |
0,968 |
0,456 |
1,424 |
2,12 |
0,905 |
0,347 |
1,253 |
2,6 |
1,34 |
1,632 |
|
Чукурова-4 |
1,045 |
0,423 |
1,468 |
2,47 |
1,084 |
0,405 |
1,489 |
2,67 |
1,18 |
1,53 |
|
Чукурова-5 |
1,006 |
0,384 |
1,390 |
2,62 |
1,073 |
0,398 |
1,471 |
2,69 |
1,26 |
1,6 |
|
Чукурова-6 |
1,049 |
0,501 |
1,55 |
2,09 |
1,099 |
0,401 |
1,5 |
2,74 |
1,07 |
1,67 |
|
Чукурова-7 |
0,787 |
0,246 |
1,033 |
3,20 |
0,943 |
0,286 |
1,229 |
3,29 |
0,93 |
1,48 |
|
Чукурова-8 |
0,986 |
0,298 |
1,284 |
3,30 |
0,963 |
0,223 |
1,186 |
4,32 |
1,23 |
1,59 |
|
Чукурова-9 |
1,023 |
0,328 |
1,351 |
3,12 |
1,108 |
0,373 |
1,481 |
2,97 |
1,37 |
1,91 |
|
Чукурова-12 |
0,815 |
0,301 |
1,116 |
2,71 |
0,791 |
0,253 |
1,044 |
3,13 |
1,08 |
1,25 |
|
Чукурова-13 |
1,405 |
0,531 |
1,936 |
2,64 |
1,176 |
0,433 |
1,609 |
2,71 |
1,53 |
1,88 |
|
Чукурова-14 |
1,219 |
0,454 |
1,673 |
2,68 |
1,387 |
0,451 |
1,838 |
3,07 |
0,87 |
1,84 |
|
Kанада-1 |
1,504 |
0,522 |
2,026 |
2,88 |
1,863 |
0,540 |
2,403 |
3,45 |
1,48 |
0,93 |
|
Kанада-4 |
0,91 |
0,331 |
1,241 |
2,74 |
1,282 |
0,441 |
1,723 |
2,91 |
0,9 |
1,91 |
|
Kанада-5 |
1,061 |
0,329 |
1,39 |
3,22 |
1,002 |
0,306 |
1,308 |
3,27 |
1,33 |
1,71 |
|
Kанада-6 |
1,416 |
0,677 |
2,093 |
2,09 |
1,239 |
0,538 |
1,777 |
2,30 |
0,72 |
1,24 |
|
Kанада-7 |
1,025 |
0,327 |
1,352 |
3,13 |
0,931 |
0,386 |
1,317 |
2,41 |
1,21 |
1,27 |
|
Уманская |
1,301 |
0,47 |
1,771 |
2,76 |
1,184 |
0,415 |
1,599 |
2,85 |
1,58 |
1,86 |
|
Антониа |
1,193 |
0,419 |
1,612 |
2,84 |
1,105 |
0,391 |
1,541 |
2,83 |
1,22 |
1,84 |
|
Aяз |
0,925 |
0,357 |
1,282 |
2.59 |
1,021 |
0,351 |
1,372 |
2,91 |
0,85 |
1,6 |
|
Aнгелика |
1,318 |
0,494 |
1,812 |
2,67 |
1,40 |
0,503 |
1,903 |
2,78 |
1,45 |
1,48 |
|
Aлекса |
1,371 |
0,506 |
1,877 |
2,71 |
1,217 |
0,434 |
1,651 |
2,80 |
1,59 |
1,59 |
|
Kиота |
1,425 |
0,497 |
1,922 |
2,87 |
1,188 |
0,419 |
1,607 |
2,83 |
1,51 |
1,6 |
Заключение
В условиях водного стресса низкий уровень дефицита воды соответствовал высокому относительному содержанию воды у образцов сортов Чукурова-3, Чукурова-7, Канада-4, Канада-6, Антониа, Ангелика. У образцов сои Чукурова-7, Чукурова-14, Канада-1, Канада-5, Канада-7, Уманская, Ангелика, Антониа, Алекса и Киото потеря воды оказалась ниже 45,0 %. Учитывая, что для устойчивых сортов в нормальных естественных условиях произрастания характерно содержание относительной воды в тканях от 70,0 – 80,0 % и выше, образцы со значениями водоудерживающей способности выше 50,0 % в условиях засухи, можно считать более устойчивыми (Канада-1, Канада-7, Антониа, Ангелика). Несмотря на относительно низкое значение водоудерживающей способности, образец Чукурова-7 также отличился засухоустойчивостью.
В условиях засухи сумма xl(a+b) относительно контроля составляла более 100 % у образцов сортов сои Чукурова-4, Чукурова-5, Чукурова-7, Чукурова-9, Чукурова-14, Канада-1, Канада-7, Аяз, Ангелика. У большинства образцов наблюдали увеличение содержания каротиноидов.