
Scientific journal
International Journal of Applied and fundamental research
ISSN 1996-3955
ИФ РИНЦ = 0,556
EFFECT OF EXOGENOUS NITRATES ON CO2/H2O EXCHANGE PARAMETERS IN KARELIAN BIRCH AND SILVER BIRCH

Видовое разнообразие лесных сообществ определяется гидрометеорологическими переменными и эдафическими условиями, а также идификационной ролью видов-лесообразователей. В условиях Северо-Запада России основными лесообразующими видами являются древесные рода Betula L. наряду с растениями родов Pinus L. и Picea L. Различные условия произрастания обуславливают высокий полиморфизм фенотипических признаков представителей рода Betula L. Большой интерес у исследователей вызывают растения березы повислой (Betula pendula Roth), особенностью которой является способность образовывать две формы: обычной березы повислой с прямослойной древесиной и карельской березы (Betula pendula Roth var. carelica (Merclin) Hamet Ahti) с узорчатой древесиной. Для карельской березы отмечают аномальное строение проводящих тканей осевых органов, которое проявляется в свилеватости структурных элементов древесины, значительном уменьшении количества сосудов и волокнистых трахеид, увеличении количества клеток древесной паренхимы [3]. Эти особенности структурной организации тканей, вероятно, должны проявляться на обменных процессах. Ранее нами было установлено увеличение степени различий водного дефицита и насыщающего содержания воды в листе обеих берез с усилением узорчатости древесины ствола карельской березы в онтогенезе [6]. Исследование минерального состава березы повислой и карельской березы в естественных условиях произрастания позволило нам выявить количественные различия содержания макроэлементов в листе и проводящих тканях деревьев исследуемых берез и больший диапазон их варьирования у карельской березы на фоне их сходной сезонной динамики [4]. В камеральных условиях нами было выявлено сходство «биологического» и «хозяйственного» оптимумов N:P:K березы повислой и карельской березы, и показаны внутривидовые особенности распределения биомассы растения в условиях разной обеспеченности среды макроэлементами [5]. Однако остается много вопросов об эколого-физиологических особенностях карельской березы с уникальной узорчатой текстурой древесины.
Вопрос о роли воздействия факторов среды на формирование узорчатой древесины карельской березы широко обсуждается в литературе [1, 3, 2]. Важную роль в комплексе внешних факторов, индуцирующих аномальный рост, отводят уровню почвенного плодородия, в частности азотного питания. В этой связи представляет интерес комплексное изучение показателей СО2 и Н2О обмена обычной березы повислой (Betula pendula var. pendula) и карельской березы (Betula pendula var. сarelica) при разной обеспеченности среды нитратами.
Материалы и методы исследования
Исследование проводили на экспериментальных участках Института леса КарНЦ РАН на территории Агробиологической станции КарНЦ РАН (N 61 °45′, Е 34 °20′). В качестве объектов исследований были выбраны 6-летние деревья обычной березы повислой (Betula pendula var. pendula) и карельской березы (Betula pendula var. сarelica) с проявившимися признаками структурных аномалий, произрастающие в полевых условиях на двух участках: 1) без внесения удобрений – контроль, 2) с внесением нитрата калия (KNO3) – опыт. В опыте под каждое дерево однократно вносили 20 л 50 мМ раствора KNO3, в контроле – соответственно, 20 л Н2О [2]. Биологическая повторность трехкратная.
Измерение устьичной проводимости (gs), интенсивности фотосинтеза (А) и транспирации (E) листа проводили в дневной динамике с 9.00 до 15.00 в июле на неотделенных листьях в средней части кроны берез с помощью портативной фотосинтетической системы Li-Cor 6400ХТ (Li-Cor Inc., США). В работе использовали стандартную листовую камеру с источником света Li-Cor 6400-02B LED (Li-Cor Inc., США). Наблюдения у всех растений проводили на сформировавшихся листьях по единой методике в листовой камере при освещенности, концентрации СО2, температуре воздуха и интенсивности потока воздуха соответственно – 1600 мкмоль м-2 с-1, 400 мкмоль СО2 моль-1, 24 °С и 500 мкмоль с-1.
Для определения других показателей водообмена со средней части кроны каждого растения отбирали по 3 облиственных побега и 10 листьев с соседних побегов. В облиственных побегах определяли водный потенциал (ψ) с помощью камеры давления Plant Moisture Vessel SKPM 1400 (Skye Instruments Ltd., Великобритания). На отрезанных листьях определяли свежую массу (wf), массу при насыщении (ws), сухую массу листьев (wd). Содержание воды в листьях (WCf) и дефицит водного насыщения (WSD) определяли по формулам: 1) WCf = (Wf – Wd) / Wd; 2) WSD = (ws – wf) / (ws – wd). Фотосинтетическую эффективность использования воды (WUE) вычисляли по отношению скоростей фотосинтеза (A) и транспирации (E), а эффективность использования азота (NUE) – по отношению скорости фотосинтеза к содержанию азота в листе [9]. Определение содержания азота (N) выполняли после мокрого озоления на оборудовании ЦКП «Аналитическая лаборатория» ИЛ КарНЦ РАН. Для обработки результатов использовали методы вариационной статистики. На графиках приведены средние арифметические значения и их стандартные ошибки. Проверку гипотез и оценку существенных различий между средними величинами осуществляли при 5 %-ном уровне значимости.
Результаты исследования и их обсуждение
В ходе проведенного эксперимента нами были установлены следующие закономерности. Сравнительный анализ показателей СО2 и Н2О обмена выявил однонаправленную реакцию березы повислой и карельской березы на внесение высоких доз нитратного азота. Нами была установлена тенденция снижения устьичной проводимости (gs), интенсивности фотосинтеза (А), транспирации (Е), водного потенциала (ψ), содержания воды (WCf) и водного дефицита (WSD) листа для обеих исследуемых форм берез при подкормке растений экзогенными нитратами (рис. 1). Так, в контроле значения gs, А, Е, Ψ, WCf и WSD листа березы повислой и карельской березы составили, соответственно, 0.29 и 0.23 моль Н2О м-2 с-1, 18.26 и 17.51 мкмоль СО2 м-2 с-1, 3.28 и 2.99 млмоль Н2О м-2 с-1, – 1.08 и -0.94 МПа, 1.36 и 1.35 гводы г-1сух.массы, 22.63 и 22.77 %. Снижение значений gs, А, Е, Ψ, WCf и WSD у березы повислой в опытной группе по сравнению с контролем составило, соответственно, 14, 8, 5, 19, 7 и 18 %, у карельской березы – 17, 6, 15, 37, 4 и 35 % соответственно. Этот факт, вероятно, обусловлен тем, что при внесении высоких доз нитратов в почву происходит увеличение концентрации почвенного раствора и, соответственно, снижение Ψ почвенного раствора. Это в свою очередь затрудняет поступление и транспорт воды и питательных веществ в системе целого растения, вследствие чего происходит снижение исследуемых показателей листа обеих форм берез. Ранее в работе Н.А. Галибиной с соавт. [2] на тех же экспериментальных объектах было показано, что внесение 50 мМ раствора нитрата калия вызвало 100-кратное увеличение нитратного азота в почве и привело к возрастанию его содержания в корнях опытных растений в 8–9 раз по сравнению с контрольными деревьями. При этом в листе берез опытной группы после внесения высокой дозы азотного удобрения нами было показано снижение содержания N по сравнению с контролем как у березы повислой, так и у карельской березы на 4 и 10 %, соответственно.
Снижение WSD листа берез, который, казалось бы, должен усиливаться при внесении высоких доз азотных удобрений, вероятно, обусловлено ограничением транспирации, контролируемой устьицами посредством уменьшения gs. Ранее было показано [10 и др.], что Betula pendula относится к видам, избегающим засуху в большей степени, чем переносящим ее, вследствие чего при недостатке воды снижение интенсивности транспирации листа происходит в результате закрытия устьиц. Кроме того, в условиях высокого содержания азота в почве ограничение устьичной проводимости и интенсивности транспирации и, соответственно, снижение фотосинтеза травянистых растений связывают с увеличением гидравлического сопротивления транспортных путей [8]. Отмечают также, что, несмотря на усиление устьичного контроля для избежания больших потерь воды на фоне увеличения обеспеченности макроэлементами, такая корректировка может быть недостаточной для предотвращения увеличения водного дефицита, вследствие чего водный потенциал станет более отрицательным [7].
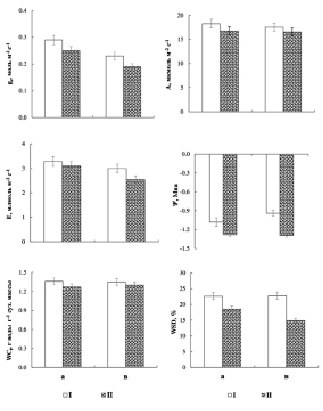
Рис. 1. Устьичная проводимость (gs), интенсивность фотосинтеза (А), интенсивность транспирации (E), водный потенциал (Ψ), содержание воды (WCf) и водный дефицит (WSD) листа березы повислой (а) и карельской березы (в) в контроле (I) и опыте (II)
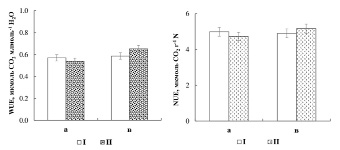
Рис. 2. Эффективность использования воды (WUE) и эффективность использования азота (NUE) листа березы повислой (а) и карельской березы (в) в контроле (I) и опыте (II)
Сопоставление коэффициентов эффективности использования воды (WUE) и азота (NUE) листа исследуемых форм берез выявило их сходство в контрольном варианте и разнонаправленные изменения в варианте с удобрениями (рис. 2). Так, значения WUE и NUE листа березы повислой и карельской березы в контроле были близки и составили, соответственно, 0.57, 0.59 мкмоль СО2 млмоль-1 Н2О и 4.99, 4.90 мкмоль СО2 г-1 N.
После внесения нитратов значения WUE и NUE листа березы повислой уменьшились на 5 % и составили 0.54 мкмоль СО2 млмоль-1 Н2О и 4.73 мкмоль СО2 г-1 N соответственно; у карельской березы, напротив, WUE и NUE увеличились на 10 и 5 % и составили, соответственно, 0.65 мкмоль СО2 млмоль-1 Н2О и 5.16 мкмоль СО2 г-1 N. Факт роста коэффициентов WUE и NUE листа карельской березы, по сравнению с березой повислой, вероятно, может свидетельствовать о более экономном расходовании влаги и более активном включении азота в метаболические процессы в условиях высокой обеспеченности почвы нитратами.
Заключение
Проведенное исследование влияния экзогенных нитратов на показатели CO2/H2O обмена карельской березы (Betula pendula Roth var. carelica) и березы повислой (Betula pendula Roth) позволило выявить некоторые внутривидовые различия. В контрольной группе растений более высокие значения gs, А, Е и Ψ листа отмечены для березы повислой по сравнению с карельской березой, и близкие значения WCf , WSD, WUE и NUE листа обеих исследуемых форм берез. При внесении высокой дозы нитратов установлено снижение всех исследуемых показателей СО2 и Н2О обмена как у березы повислой, так и у карельской березы. Однако большие изменения значений Е, Ψ, WSD и WUE отмечены для карельской березы по сравнению с березой повислой, что может свидетельствовать о различиях в напряженности водообмена исследуемых берез вследствие внутривидовых особенностей структуры проводящих тканей ствола дерева.
Работа выполнена в рамках государственного задания ИЛ КарНЦ РАН (проект № 0220-2014-0001).
Библиографическая ссылка
Придача В.Б., Новичонок Е.В., Сазонова Т.А. ВЛИЯНИЕ ЭКЗОГЕННЫХ НИТРАТОВ НА ПОКАЗАТЕЛИ CO2/H2O ОБМЕНА КАРЕЛЬСКОЙ БЕРЕЗЫ И БЕРЕЗЫ ПОВИСЛОЙ // Международный журнал прикладных и фундаментальных исследований. 2016. № 10-4. С. 591-594;URL: https://applied-research.ru/en/article/view?id=10395 (дата обращения: 19.06.2026).