
Scientific journal
International Journal of Applied and fundamental research
ISSN 1996-3955
ИФ РИНЦ = 0,556
INFLUENCE OF VRN-B1 ALLELES ON THE PHASE DURATION OF DEVELOPMENT OF SUBSTITUTED AND ISOGENIC WHEAT LINES UNDER NATURAL LONG DAY

Важным фактором, определяющим адаптацию растений (в том числе, мягкой пшеницы) к условиям окружающей среды, является продолжительность фаз развития, особенно сроки наступления колошения [19, 23]. Фенологические фазы развития, определяющие жизненный цикл пшеницы, предполагают определение всходов, третьего листа, кущения, первого узла, выхода в трубку, флагового листа, колошения и спелости. При этом установлено, что в своем развитии пшеница до фазы «выход в трубку» находится в вегетативном периоде развития, от начала колошения до конца цветения – в генеративном, и от первой стадии созревания до полной спелости – в репродуктивном периоде [3]. Различные фазы определяют формирование определенных количественных показателей продуктивности [16]. Так, более продолжительный период «всходы–кущение» приводит к увеличению вегетативной массы растений (по Ф.М. Куперман [3] – I–II этапы органогенеза) [20]. Продолжительность фазы «выход в трубку» (IV–VII этапы органогенеза) весьма важна, поскольку в этот период происходит формирование фертильных цветков, определяющих будущее число зерен в колосе [7, 18]. Во время периода «колошение–созревание» (IX–XII этапы органогенеза) происходят налив и увеличение массы зерна [17]. Показано, что время наступления и длительность различных фаз развития контролируются генетически [2].
Гены чувствительности к яровизации (Vrn) и фотопериоду (Ppd) и гены раннеспелости (Eps) влияют на продолжительность жизненного цикла мягкой пшеницы. При этом гены Vrn определяют до 75 % различий по длине вегетационного периода [9]. У мягкой пшеницы известно пять главных генов Vrn [11] – Vrn-A1, Vrn-B1, Vrn-D1, Vrn-D4, Vrn-B3, доминантный аллель хотя бы в одном из которых определяет яровой тип развития, а рецессивные по всем этим генам – озимый тип развития [12]. За счет множественных аллелей доминантных локусов Vrn-1 возможно увеличение разнообразия мягкой пшеницы по длине вегетационного периода [5, 15, 22]. У двух генов – Vrn-A1 и Vrn-B1 обнаружено наличие множественных аллелей, которые влияют на различия во времени колошения [4, 6]. При проведении данных исследований эти аллели находились в разной генотипической среде, после чего был проведен анализ времени колошения и реакции на яровизацию различных сортов и линий.
Большую роль в изучении генов Vrn сыграли изогенные линии по доминантным генам Vrn, созданные на основе сорта Triple Dirk [11, 12]. Сам сорт Triple Dirk имеет два доминантных гена – Vrn-A1 и Vrn-B1. Изогенная линия Triple Dirk D имеет доминантный ген Vrn-A1, линия Triple Dirk B – ген Vrn-B1, линия Triple Dirk E – ген Vrn-D1, линия Triple Dirk F – ген Vrn-D4. Донорами генов Vrn-A1 и Vrn-B1 послужил сорт Triple Dirk, донором гена Vrn-D1 – сорт Loro, а донором гена Vrn-D4 – сорт Gabo. У изогенной линии Triple Dirk C все четыре гена Vrn – рецессивные. Донором рецессивных аллелей vrn послужил озимый сорт Winter Minflor [14, 21].
По фенотипическому проявлению доминантные гены Vrn в изогенных линиях сорта Triple Dirk располагаются в следующем порядке: Vrn-A1 > Vrn-D1 > Vrn-D4 ≥ Vrn-B1: изогенная линия Triple Dirk D с геном Vrn-A1 является самой раннеспелой, а линия Triple Dirk B с геном Vrn-B1 – самой позднеспелой; линия с геном Vrn-D4 не отличается достоверно по времени колошения от линии с геном Vrn-B1 [10].
Для изучения множественного аллелизма доминантного гена Vrn-B1 были созданы замещённые лини сорта Sava по хромосоме 5В и изогенные линии сорта Безостая 1 [14] (рис. 1). Донорами хромосом 5В с доминантными аллелями Vrn-B1c и Vrn-B1a и самих аллелей Vrn-B1c и Vrn-B1a для этих линий послужили замещённые линии С29/М808 5А и ДмII/М808 5А, полученные Майстренко (1992). При создании замещённых линий сорта Sava по хромосоме 5В моносомная линия сорта Sava по хромосоме 5В, предоставленная Э. Ворладном, была скрещена с замещёнными линиями С29(М808 5А) и Дм2 (М808 5А). Цитологически выделенные моносомные растения с хромосомой 5В от линий С29 (М808 5А) или Дм II (М808 5А) самоопыляли. С цитологически выделенными дисомными растениями проводили семь поколений беккроссов моносомной линией сорта Sava по хромосоме 5В для восстановления генотипа озимого сорта Sava, с цитологическим выделением после каждого беккросса моносомных и дисомных растений. В результате замещённые линии в генетическом фоне озимого сорта Sava, рецессивного по генам Vrn-1, содержали хромосомы 5В от яровых сортов Саратовская 29 и Диамант 2 с доминантными аллелями Vrn-B1c и Vrn-B1a [14]. При создании изогенных линий сорта Безостая 1 озимый сорт Безостая 1 скрещивали с линиями С29 (М808 5А) и Дм2 (М808 5А) и полученные растения беккроссировали 8 раз озимым сортом Безостая 1 для восстановления генотипа этого озимого сорта. В первом и третьем поколениях беккросса при помощи SSR-маркёров были отобраны растения, имевшие наименьшую вставку донорской хромосомы 5В, содержащей аллели гена Vrn-B1, которые использовали в следующих беккроссах. Полученные в результате изогенные линии в генетическом фоне озимого сорта Безостая 1, рецессивного по генам Vrn-1, имели аллели Vrn-B1c и Vrn-B1a от яровых сортов Саратовская 29 и Диамант 2 [14].
Изогенные линии сорта Безостая 1 с доминантными генами Vrn-A1 и Vrn-D4 были получены по аналогичной схеме (Ефремова Т. Т., неопубл.). Донором доминантного гена Vrn-A1 для изогенной линии i:Без1Vrn-A1 послужила изогенная линия Triple Dirk D [12], а донором доминантного гена Vrn-D4 для изогенной линии i:Б1Vrn-D4 – образец T. sphaerococcum Persiv. к-5498 из коллекции ВИР.
На время колошения, а также на связанные с ним компоненты урожайности гены Vrn влияют не только по отдельности, но и в комбинации друг с другом [19]. Так, сорта с двумя доминантными генами Vrn выколашиваются раньше сортов с одним геном Vrn, а сорта с тремя доминантными генами Vrn являются самыми раннеспелыми. Однако сорта с тремя доминантными генами Vrn имеют наименьшую продуктивность, так как у них снижены показатели компонентов урожайности и масса 1000 зёрен. Самыми высокоурожайными являются сорта с одним геном Vrn-A1 или двумя генами Vrn-A1 и Vrn-B1: они имеют хорошие показатели продуктивности, а раннеспелость позволяет им в полной мере пройти налив зерна. Сорта с доминантным геном Vrn-B1 или комбинацией Vrn-B1 Vrn-D1 имеют высокие показатели признаков продуктивности, но позднеспелость не позволяет им в достаточной степени пройти налив зерна, поэтому по продуктивности они уступают сортам с геном Vrn-A1 и двумя генами Vrn-A1 и Vrn-B1. Носители доминантного гена Vrn-D1, одного, или в комбинации с генами Vrn-A1 или Vrn-B1, по продуктивности не превосходят перечисленные сорта, но имеют преимущество при выращивании растений в регионах со стрессовыми условиями в период налива зерна, такими, как засуха и высокая температура [13].
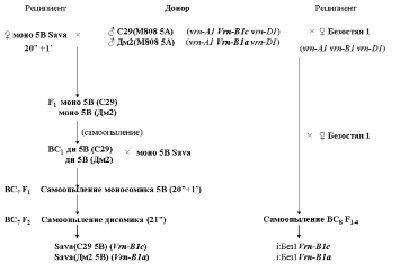
Рис. 1. Схемы получения замещённых линий озимого сорта Sava по хромосоме 5В и изогенных линий озимого сорта Безостая 1 с аллелями Vrn-B1c и Vrn-B1a [14]
Для выяснения влияния разных аллелей на скороспелость и агрономические признаки необходимо было создать изогенные линии по двум аллелям гена Vrn-В1. Ими оказались две замещенные линии на основе озимого сорта Sava хромосоме 5В и две изогенные линии на основе озимого сорта Безостая 1 (Б1), обе несут аллели Vrn-B1c и Vrn-B1a от сортов Саратовская 29 (С29) и Диамант II (ДмII) [14]. Исследование выявило, что замещенные и изогенные линии с аллелем Vrn-B1c выколашиваются раньше, чем линии с аллелем Vrn-B1a. В то же время, на яровизацию сильнее реагируют линии с аллелем Vrn-B1a, чем с аллелем Vrn-B1c. Кроме того оказалось, что замещенные и изогенные линии с аллелем Vrn-B1с характеризуются достоверно меньшей продолжительностью периодов «кущение – первый узел» и «выход в трубку – колошение» и, следовательно, выколашиваются раньше [21]. Однако эти результаты были получены при выращивании в условиях теплицы, а в полевых условиях такие исследования не проводились. Поэтому остался ряд вопросов, связанных с поведением вышеописанных сортов и линий пшеницы в естественных условиях. Данные обстоятельства определили актуальность настоящего исследования.
Целью работы являлось изучение продолжительности фаз развития у замещённых и изогенных линий мягкой пшеницы с разными аллелями гена VRN-B1 при естественном длинном дне в условиях лесостепной зоны Приобья Новосибирской области.
Материалы и методы исследования
Объектом исследования послужили замещённые линии озимого сорта Sava по хромосоме 5В и изогенные линии озимого сорта Безостая 1 [14], а также другие линии и сорта из коллекции лаборатории хромосомной инженерии злаков ИЦиГ СО РАН (таблица 1).
У растений отмечались следующие фазы развития: всходы, появление, 3–го листа, кущение, появление первого узла, выход в трубку, колошение, созревание. Даты наступления фаз развития отмечались у каждого растения индивидуально и высчитывалось среднее значение. Каждой линии или сорта выращивалось от 10 до 40 растений.
Методика проведения эксперимента
Опыт проводился в течение полугода. Предшественником служил пар. Перед посевом было произведено внесение удобрения (Азофоска – N60P60K60) и боронование, а также разбивка и маркировка поля на яруса.
Схема посева: 50 см х 70 см (50 – ширина яруса, 70 – ширина дорожки). Расстояние между рядками – 25 см. Норма высева на погонный метр – 20–25 семян. Глубина заделки семян составляла 3–4 см. Дата посева – 13 мая. После посева было произведено уплотнение почвы и нарушение верхнего слоя с целью уменьшения испарения влаги из почвы. Защитное мероприятие по борьбе со шведской мухой – покрытие агрилом (нетканый укрывной материал).
Таблица 1
Материал исследования
|
Линия, сорт |
Гаплоидный генотип по генам Vrn |
Донор доминантного гена Vrn |
Литературный источник |
|
Sava(С29 5В) |
vrn-A1 Vrn-B1c vrn-D1 |
Саратовская 29 |
– |
|
Sava(ДмII 5В) |
vrn-A1 Vrn-B1a vrn-D1 |
Диамант II |
– |
|
i:Без1Vrn-B1c |
vrn-A1 Vrn-B1c vrn-D1 |
Саратовская 29 |
Efremova et al., 2011, Shcherban et al., 2012a |
|
i:Без1Vrn-B1a |
vrn-A1 Vrn-B1a vrn-D1 |
Диамант II |
– |
|
i:Без1Vrn-A1 |
Vrn-A1 vrn-B1 vrn-D1 |
Triple Dirk D |
Ефремова (неопуб.) |
|
i:Без1Vrn-D4 |
vrn-A1 vrn-B1 vrn-D1 vrn-D4 |
T. sphaerococcum Persiv. к-5498 |
– |
|
С29/М808 5А |
vrn-A1 Vrn-B1c vrn-D1 |
Саратовская 29 |
Майстренко, 1992 |
|
ДмII/М808 5А |
vrn-A1 Vrn-B1a vrn-D1 |
Диамант 2 |
– |
|
Саратовская 29 |
Vrn-A1 Vrn-B1c vrn-D1 |
– |
|
|
Диамант II |
Vrn-A1 Vrn-B1a vrn-D1 |
– |
|
|
Mara |
vrn-A1 Vrn-B1a vrn-D1 |
Shcherban et al., 2012a |
|
|
Federation |
vrn-A1 Vrn-B1c vrn-D1 |
– |
|
|
Омская 9 |
vrn-A1 Vrn-B1c vrn-D1 |
– |
|
|
Мильтурум 553 |
vrn-A1 Vrn-B1c vrn-D1 |
||
|
Мильтурум 321 |
vrn-A1 Vrn-B1c vrn-D1 |
||
|
Tr. sphaerococcum |
Vrn-A1Vrn-B1а vrn-D1 |
Гончаров, 2012 |
|
|
Цезиум 94 |
Vrn-A1Vrn-B1с vrn-D1 |
||
|
Gabo |
vrn-A1 Vrn-B1 vrn-D1 Vrn-D4 |
Pugsley, 1972 |
Во время наступления фазы всходов, кущения, выход в трубку была произведена обработка растений препаратом «Актеллик» (10 мл на 10 л воды) от комплекса насекомых: шведская муха, крестоцветная блошка. Время от времени на поле производилось рыхление, с целью проникания воздуха в почву и прополка междурядий, дорожек и периметра поля, с целью уборки сорняков. После наступления фазы флаговый лист у всех растений на поле было произведено крепление этикеток с порядковым номером на каждое первое растение в рядке, для облегчения дальнейшей работы.
Начало даты уборки – 19 августа. Разметка поля, посев, уход за растениями в течение вегетации и уборка производились вручную. Обмолот экспериментального материала производился с помощью электрической колосковой молотилки МКС-1М.
Даты наступления и продолжительности каждой фазы развития для каждой отдельной линии заносились в журнал наблюдений, а затем подверглись статистическому анализу.
Условия проведения эксперимента
Институт цитологии и генетики СО РАН расположен в лесостепной зоне, вблизи города Новосибирска, на берегу Обского водохранилища. Особенности климата Новосибирской области связаны с её положением в центре материка и характером рельефа этого региона. Ярко выраженный континентальный климат – это суровая продолжительная зима и короткое, жаркое, часто засушливое лето. Главное в климате – это высокий приход лучистой энергии и нехватка тепла. Продолжительность теплового периода с температурами выше 0°С составляет 185 – 195 дней. Осадков выпадает 300–350 мм в год. Наибольшее количество осадков обычно выпадает в июле – 60–80 мм. На зимний период приходится 17–20 % годовой суммы осадков. Периодические засухи в весенне-летний период отрицательно сказываются на жизнедеятельности растений.
Почвы серые лесные, развивающиеся на породах, контактирующих с отложениями древней террасы реки Оби. По механическому составу почвы среднесуглинистые, с большим количеством фракции крупной пыли, бесструктурные, со слабой воздухо- и водопроницаемостью, быстрой осадкой и уплотнением после обработки, склонные к заплыванию и образованию корки. Реакция слабокислая. Агрохимический анализ показал, что почва содержит относительно небольшое количество гумуса (3,85 %). Близость леса защищает от ветровой эрозии и выветривания снежного покрова.
Температура в год исследования практически не отличалась от среднемноголетней нормы. В 2016 г. количество осадков за вегетационный период выпало ниже среднемноголетнего, самым засушливым месяцем был август. В июле осадков выпало чуть больше нормы. Осадков за вегетационный период выпало 167 мм (на 53 мм меньше нормы).
Таблица 2
Температура воздуха и осадки за вегетационный период 2016 года по ГМС «Огурцово»
|
Температура |
Осадки |
|||||||||||
|
Месяц |
I декада |
II декада |
III декада |
Ср. месяч. |
Норма |
Откл. от нормы |
I декада |
II декада |
III декада |
Сумма за месяц |
Норма |
% от нормы |
|
Май |
6,5 |
8,7 |
15,8 |
10,3 |
10,9 |
-0,6 |
7 |
11 |
14 |
32 |
37 |
86,5 |
|
Июнь |
17,5 |
20,5 |
21,1 |
19,7 |
16,9 |
2,8 |
0 |
28 |
10 |
38 |
55 |
69 |
|
Июль |
19,9 |
21 |
19,8 |
20,2 |
19,4 |
0,8 |
47 |
16 |
14 |
77 |
61 |
126,2 |
|
Август |
18,2 |
18 |
15,9 |
17,4 |
16,2 |
1,2 |
12 |
0 |
8 |
20 |
67 |
29,9 |
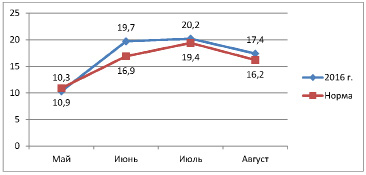
Рис. 2. Температура за вегетационный период
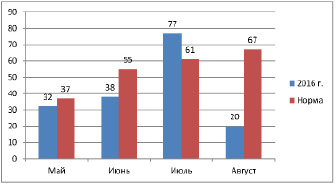
Рис. 3. Осадки за вегетационный период
Статистическая обработка данных выполнена с использованием программы Excel, версия: 14.0.7128.5000 (32–разрядная). Определялся характер распределения количественных признаков методом Колмогорова-Смирнова. В случае нормального распределения вычислялось среднее значение и стандартная ошибка среднего. Достоверность различия показателей оценивали по критериям Стьюдента, Пирсона (при нормальном распределении), в случаях отклонения распределения от нормального использовались непараметрические критерии (U-критерий Манна-Уитни, Колмогорова-Смирнова). Во всех процедурах статистического анализа критический уровень значимости нулевой гипотезы (p) принимался равным 0,05.
Результаты исследования и их обсуждение
Продолжительность фаз развития у сортов мягкой пшеницы, изогенных и замещенных линий с разными аллелями локуса Vrn в условиях лесостепной зоны Новосибирской обл. (посев 13 мая 2016 г.) представлен в таблице 3.
Анализ данных, представленных в таблице 3, выявил, что наиболее скороспелыми были сорта и линии с доминантным геном Vrn-A1, которые выколашивались в интервале 38–41 дней: Саратовская29, Диамант II, Gabo, Мильтурум553. Полученные результаты согласуются с данными по выращиванию изогенной линии Vrn-A1 в условиях длинного дня, где данная линия оказалась самой раннеспелой, она выколосилась на 41 день, на 11 дней раньше, чем изогенная линия с аллелем Vrn-B1с (Р>0,999) [21]. Оказалось, что сорта и линии с одним доминантным геном Vrn-B1 более позднеспелые и выколашиваются в интервале 44–52 дней: C29/5A М808, ДмII/5АМ808, Омская 9, Federation, Mara, Sava/5B C29, Sava/5B ДмII, T. sphaeroccocum, Мильтурум 321, Цезиум 94. Более длительные сроки колошения изогенных линий с аллелем Vrn-B1а были установлены при выращивании в условиях теплицы (на 69-й день от всходов, Р>0,999 [21].
Вместе с тем, выявилось, что замещенные и изогенные линии с аллелем Vrn-B1c выколашиваются раньше, чем линии с аллелем Vrn-B1a на 2–3 дня. В то же время, в условиях длинного дня, по данным Емцевой М.В. и соавт., различие во времени колошения между изогенными линиями соответствовало 17 дням [21].
Длительность периода «всходы – кущение» у изогенных и замещённых линий практически не различалась. Анализ продолжительности фаз развития установил, что основные различия по прохождению фаз развития у линий с разными аллелями локуса Vrn начинаются с фазы «кущение-выход в трубку». Период «кущение-выход в трубку» является критическим и определяет продолжительность всего вегетационного периода от всходов до колошения и созревания растений [21].
Таблица 3
Продолжительность фаз развития замещённых и изогенных линий мягкой пшеницы с разными аллелями локусов VRN1
|
Линия, сорт |
Число изученных растений |
Продолжительность фаз развития, сутки |
||||||
|
Всходы – 3-й лист |
Всходы – кущение |
Всходы –1-й узел |
Всходы – выход в трубку |
Всходы –флаговый лист |
Всходы – колошение |
Всходы-восковая спелость |
||
|
i:Б1 Vrn-Blc |
39 |
5,10±0,64 |
10,28±1,88 |
27,51±1,17 |
30,95±1,37 |
35,18±1,65 |
44,46±1,55 |
88,82±0,39 |
|
i:Б1 Vrn-Bla |
18 |
5,18±0,88 |
11,07±1,71 |
28,33±1,62 |
32,33±2,20 |
36,78±1,76 |
46,92±1,98 |
88,26±0,45 |
|
i:Б1 Vrn-D4 |
35 |
5,57±0,96 |
11,42±1,89 |
28,11±1,13 |
32,14±1,14 |
38,26±2,54 |
48,79±2,27 |
94,65±0,77 |
|
i:Б1 Vrn-Ala |
26 |
5,33±0,68 |
10,88±1,37 |
23,88±1,24 |
27,50±1,24 |
31,73±2,44 |
41,81±2,81 |
88,54±0,51 |
|
ДмII/5АМ808 |
26 |
5,92±1,72 |
11,38±1,96 |
31,61±1,02 |
35,73±1,34 |
41,88±1,27 |
50,80±1,15 |
92,64±0,49 |
|
C29/5A М808 |
33 |
5,70±0,77 |
10,61±1,32 |
25,39±1,98 |
29,24±1,73 |
36,10±1,53 |
44,72±2,00 |
88,61±0,50 |
|
Sava/5B ДмII |
32 |
5,26±1,44 |
10,65±2,25 |
34,19±1,69 |
38,53±2,00 |
43,75±3,40 |
51,32±2,90 |
94.84±0,37 |
|
Sava/5B C29 |
17 |
5,00±1,09 |
10,75±2,10 |
32,56±2,28 |
36,56±2,70 |
41,06±3,84 |
49,87±3,26 |
94,62±0,50 |
|
Мильтурум 553 |
13 |
5,57±1,65 |
9,93±2,52 |
21,93±2,43 |
26,38±2,29 |
31,77±2,77 |
39,77±2,59 |
88,00±0,00 |
|
Омская 9 |
14 |
5,00±0,00 |
10,86±1,35 |
24,43±2,41 |
28,86±0,28 |
34,64±2,17 |
44,00±2,63 |
88,00±0,00 |
|
Цезиум 94 |
13 |
5,14±0,53 |
10,23±1,64 |
24,15±2,11 |
28,38±1,94 |
35,00±2,16 |
44,46±2,33 |
88,00±0,00 |
|
Мильтурум 321 |
8 |
5,33±1,00 |
9,67±1,41 |
22,25±2,12 |
26,87±2,17 |
32,87±5,54 |
41,50±4,67 |
88,00±0,00 |
|
T. sphaeroccocum |
24 |
5,79±1,25 |
11,17±2,26 |
31,67±1,81 |
35,87±2,25 |
40,79±3,55 |
48,46±4,23 |
94,17±1,01 |
|
Mara |
8 |
6,55±1,94 |
13,62±1,30 |
29,25±1,75 |
33,62±1,99 |
36,87±3,23 |
44,71±3,20 |
87,57±1,13 |
|
Federation |
13 |
5,07±1,54 |
10,08±1,93 |
32,61±2,18 |
37,15±1,52 |
42,46±1,81 |
52,67±3,58 |
87,75±0,87 |
|
Gаbo |
26 |
6,58±1,63 |
11,15±1,62 |
22,81±1,30 |
25,96±1,11 |
30,36±1,41 |
38,44±1,87 |
88,68±0,48 |
|
ДиамантII (ДмII) |
25 |
5,76±0,97 |
9,56±2,24 |
20,36±1,22 |
24,48±0,96 |
28,33±1,27 |
38,12±1,94 |
88,46±1,06 |
|
Саратовская 29 (С29) |
18 |
9,52±1,83 |
13,61±0,98 |
20,55±1,79 |
24,33±1,37 |
30,50±3,88 |
38,55±2,59 |
88,44±0,51 |
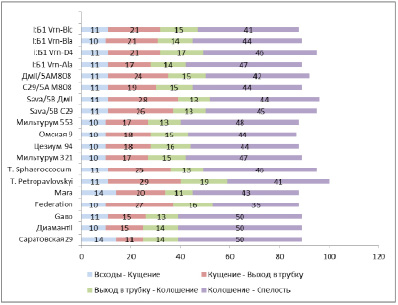
Рис. 4. Длительность фаз развития у изогенных, замещённых линий и сортов мягкой пшеницы при выращивании в условиях естественного дня (май – август 2016 г.)
Установлено, что замещённые, изогенные линии и сорта с аллелем Vrn-A1 имеют меньшую длину фазы «кущение» по сравнению с линиями – носителями аллелей Vrn-B1c и Vrn-B1a, и поэтому выколашиваются раньше. Кроме эффекта аллелей Vrn-B1c и Vrn-B1a наблюдается также влияние генетического фона сортов Sava и Безостая 1. Разница в начале фаз развития от кущения до колошения между линиями с аллелями Vrn-B1c и Vrn-B1a в генотипе сорта Sava является большей, чем в генотипе сорта Безостая 1. Замещенные линии сорта Sava были более позднеспелыми, чем изогенные линии по сорту Безостая 1. Исследование Емцевой М.В. и соавт. показало, что при отсутствии яровизации и при длинном дне замещенные и изогенные линии с аллелем Vrn-B1c имеют меньшую длину фазы «кущение» по сравнению с линиями – носителями аллеля Vrn-B1а и поэтому выколашиваются раньше [21].
Известно, что при длинном дне и неблагоприятных условиях (засуха, высокие температуры и др.) фазы развития и вегетационный период проходят быстрее, при этом закладывается меньше элементов продуктивности, а при благоприятных условиях и коротком дне развитие задерживается на III этапе органогенеза, при этом закладывается больше элементов продуктивности [1, 10].
Анализ продуктивности замещённых и изогенных линий с разными аллелями локуса Vrn в условиях лесостепной зоны Новосибирской обл. (посев 13 мая 2016 г.) (таблица 4) показал, что наиболее продуктивными по количеству зёрен, массе зерна со всего растения оказались линии i:Б1 Vrn-Bla и i:Б1 Vrn-Ala (p<0,001–0,05). Эти же линии преобладали по массе зерна, числу зерен с остальных колосьев. Линия i:Б1 Vrn-Bla лидировала и по массе зерна с колоса, однако, второй по данному параметру оказалась линия I:Б1 Vrn-Blc, заметно превосходя i:Б1 Vrn-Ala. Число зерен в колосе оказалось достоверно больше в линиях Sava (С29 5В), i:Б1 Vrn-Bl, по количеству колосков в колосе на первых позициях были линии Sava (С29 5В) и Sava (ДмII 5В). Установлено, что наибольшей длина растения с колосом была у линий I:Б1 Vrn-Bla и I:Б1 Vrn-Blc, а длина колоса оказалась максимальной в линиях Sava (С29 5В) и Sava (ДмII 5В).
Суммируя все параметры, можно сделать вывод, что наиболее продуктивными оказались линии i:Б1 Vrn-Bla и i:Б1 Vrn-Ala по сравнению с другими, принявшими участие в эксперименте.
Таблица 4
Продуктивность замещённых и изогенных линий с разными аллелями локуса Vrn в условиях лесостепной зоны Новосибирской области (посев 13 мая 2016 г.).
|
Линия, сорт |
Длина растения с колосом |
Длина колоса |
Число колосков в колосе |
Число зёрен в колосе |
Масса зерна с колоса |
Число продуктивных побегов без главного |
Число зёрен с остальных колосьев |
Масса зерна с остальных колосьев |
Число зёрен со всех ко-лосьев |
Масса зерна со всех колосьев |
Число продуктивных побегов |
|
I:Б1 Vrn-Bla |
72,44+3,81 |
8,54+0,48 |
15,89+1,08 ** |
37,00+ 5,21 |
1,41+ 0,26 |
9,00+ 2,75 |
236,22+ 72,51 |
8,81+2,75 |
273,22+ 74,07 |
10,22+2,80 |
10,00+2,75 |
|
I:Б1 Vrn-Blc |
70,42+4,54 |
8,90+0,53 |
18,04+1,52 |
32,46+ 8,95 |
1,14+ 0,40 ** |
8,12+ 3,97 |
206,58+ 95,88 |
7,06+3,31 т1 |
239,04+ 98,53 |
8,19+3,46 т1 |
9,12+3,97 |
|
I:Б1 Vrn-Ala |
64,73+5,44** |
8,44+0,55 |
15,08+1,85 ^^ |
33,20+ 9,05 |
1,02+ 0,42 т2 |
9,40+ 4,19 |
242,96+ 135,51 |
7,49+4,49 |
276,16+ 141,66 |
8,51+4,70 |
10,40+4,19 |
|
i:Б1 Vrn-D4 |
69,25+3,30 * $ |
7,74+0,51 *** $$ |
17,22+1,53 ** $$ |
31,52+ 6,52 * |
0,97+ 0,31 *** ^ |
9,74+ 5,09 |
232,63+ 116,51 |
7,02+3,47 |
264,15+ 117,71 |
7,98+3,54 |
10,74+5,09 |
|
Sava (С29 5В) |
60,13+3,25 $ |
9,76+0,57 |
21,20+2,34 ^^^ |
38,60+ 10,67 # |
0,99+ 0,32 ** |
8,40+ 6,93 |
213,13+ 171,08 |
5,14+4,06 * |
251,73+ 175,30 |
6,13+4,23 * |
9,40+6,93 |
|
Sava (ДмII 5В) |
69,00+2,85 * $ |
9,23+0,61 $ @@ т1 |
19,54+2,15 *** # @ |
35,68+ 7,91 т4 |
0,92+ 0,24 *** ^ |
8,32+ 3,14 |
214,23+ 88,64 |
5,55+2,55 * |
249,91+ 94,48 |
6,47+2,73 * |
9,32+3,14 |
Примечания:
* – достоверность отличия от I:Б1 Vrn-Bla, где: * – p<0,05;** – p<0,01;*** – p<0,001;
^ – достоверность отличия от I:Б1 Vrn-Blc, где: ^ – p<0,05; ^^ – p<0,01;^^^ – p<0,001;
$ – достоверность отличия от I:Б1 Vrn-Ala, где: $ – p<0,05; $$ – p<0,01; $$$- p<0,001;
# – достоверность отличия от i:Б1 Vrn-D4, где: # – p<0,05; ## – p<0,01; ### – p<0,001;
@ – достоверность отличия от Saval5B Cap29, где: @ – p<0,05;@@ – p<0,01; @@@ – p<0,001;
т – тенденция к наличию различий.
Заключение
В полевых условиях разница во времени колошения замещённых и изогенных линий с аллелями Vrn-B1a и Vrn-B1с составила 2–3 дня. При этом замещенные и изогенные линии с аллелем Vrn-B1c выколашиваются раньше, чем линии с аллелем Vrn-B1a. В целом замещенные линии сорта Sava были более позднеспелыми, чем изогенные линии по сорту Безостая 1. Наиболее скороспелыми были сорта и линии с доминантным геном Vrn-A1, которые выколашивались в интервале 38–41 дней. Замещённые, изогенные линии и сорта с аллелем Vrn-A1 имеют меньшую длину фазы «кущение» по сравнению с линиями – носителями аллелей Vrn-B1c и Vrn-B1a.
В полевых условиях по числу и массе зёрен со всего растения самыми продуктивными оказались изогенные линии i:Б1 Vrn-B1a и i:Б1 Vrn-A1a. Эти же линии преобладали по массе зерна, числу зерен с остальных колосьев; линия i:Б1 Vrn-Bla лидировала и по массе зерна с колоса. Остальные линии и сорта оказались достоверно ниже по продуктивности.
Таким образов, в ходе проведения эксперимента было проанализировано поведение замещённых и изогенных линий мягкой пшеницы с разными аллелями гена VRN-B1 в естественных условиях в условиях лесостепной зоны Приобья Новосибирской области, установлены самые продуктивные изогенные линии i:Б1 Vrn-B1a и i:Б1 Vrn-A1a.
Библиографическая ссылка
Кручинина Ю.В., Ефремова Т.Т., Чуманова Е.В., Попова О.М., Арбузова В.С., Першина Л.А. ВЛИЯНИЕ АЛЛЕЛЕЙ VRN-B1 НА ПРОДОЛЖИТЕЛЬНОСТЬ ФАЗ РАЗВИТИЯ ЗАМЕЩЁННЫХ И ИЗОГЕННЫХ ЛИНИЙ МЯГКОЙ ПШЕНИЦЫ ПРИ ЕСТЕСТВЕННОМ ДЛИННОМ ДНЕ // Международный журнал прикладных и фундаментальных исследований. 2017. № 1-2. С. 278-286;URL: https://applied-research.ru/en/article/view?id=11184 (дата обращения: 03.07.2026).