
Scientific journal
International Journal of Applied and fundamental research
ISSN 1996-3955
ИФ РИНЦ = 0,556
CHANGE OF MICROBIOLOGICAL SYSTEM PARAMETERS OF IRON-, MANGANESE- AND PHOSPHATE-REDUCTION OF BACTERIA AT DESTRUCTION OF DIATOMITE, ZEOLITE AND BENTONITE CLAY

Почвоведение одним из факторов почвообразования считает участие микробиологических форм жизни одним из ведущих факторов почвообразования – от первичного разрушения образующих почвы пород и вплоть до формирования самостоятельных генетических горизонтов почвенного профиля с последующей эволюцией почвенного покрова [3, 9, 10]. Однако в настоящее время в прикладных исследованиях биологии почв имеется пробел о функциональных особенностях биохимического взаимодействия вещества используемых удобрений и почвенных кондиционеров с деструкционной функцией обитающих в почве бактерий, которые принимают в этих процессах непосредственное участие [2, 5]. В особенности такая недостаточность имеется в части эффектов от прямого биохимического воздействия почвенных железо-, марганец- и фосфатредуцирующих бактерий на вещество природных кремнийсодержащих материалов [5].
Фосфатазы (фосфогидролазы, фосфорилазы – НКФ 3.1.3.17) – достаточно обширная группа ферментов, относящихся к классу гидролаз, биохимическое действие которых направлено на гидролиз разнообразных фосфорорганических соединений по фосфоэфирным связям. Данный ферментный комплекс выделяется большинством почвообитающих бактерий, как литотрофного, так и органотрофного типов питания. В результате данных процессов происходит отщепление остатков фосфорной кислоты, и фосфор органических радикалов переходит в свободное состояние. В почвах обнаружены различные фосфогидролазы: группа кислых и щелочных фосфатаз, гидролизующих моноэфиры фосфорной кислоты (глицерофосфаты, сахарофосфаты и т.д.); фитазы – особая специфическая группа, отщепляющая остатки фосфорной кислоты от фитина; группа нуклеаз (дезоксирибонуклеазы, рибонуклеазы), которые катализируют реакции деполимеризации нуклеиновых кислот [7, 8].
В связи с этим возникает необходимость в изучении прямого (контактного) взаимодействия различных фосфатредуцирующих бактерий почвы с природными высококремнистыми материалами, используемых в современной практике агрономического почвоведения.
Цель исследования
Целью исследования в настоящей работе явилось изучение динамики свободной кислотности, общей минерализации культуральной жидкости, а также численности клеток накопительной природной культуры двух групп фосфатредуцирующих бактерий (литотрофного и органотрофного типов питания), а также железо- и марганец-окисляющих бактерий дерново-подзолистой легкосуглинистой почвы Нижегородской области, при биохимической деструкции природных высококремнистых материалов, используемых в наших полевых исследованиях.
Материалы и методы исследования
Исследование было проведено в 2017 году на базе экологической и биотехнологической лабораторий ФГБОУ ВО «НГПУ им. К. Минина», оно представляло собой серию микробиологических постановочных опытов с высококремнистыми материалами различного генезиса (диатомит Инзенского, цеолит Хотынецкого и бентонитовая глина Зырянского месторождения). Породы подвергались бактериальной деструкции культурами фосфатредуцирующих, а также Fe- и Mn-окисляющих бактерий.
Культуру фосфатредуцентов-литотрофов получали на жидком питательном агаре Муромцева (АМУР) в виде его засева навеской почвы и семи суточного выращивания микробомассы при +26 °С. Культуру фосфатредуцентов-органотрофов получали на питательном агаре Менкиной (АМЕН), комплекса железо-окисляющих бактерий – на жидком агаре Бромфильда (АБ), а марганец-окисляющих – на жидком агаре Крумбайна (АК) [1, 4, 6].
Затем производился засев кремнийсодержащих материалов полученными суспензиями бактерий, после чего производилось 30-суточное культивирование систем «культура – порода». Через определенное время производилось определение жизнеспособности культуральной жидкости в виде численности живых клеток бактерий с помощью люминесцентной микроскопии с акридином оранжевым [8] на микроскопе «БиоТех-330-LED2-Tr», а также определение свободной кислотности (водородного показателя рН) микробных суспензий без гомогенизирования бактериальных клеток потенциометрическим методом с помощью рН-метра МАРК-903 и определение общей минерализации бактериальной суспензии кондуктометрическим методом с помощью кондуктометра DIST-3 HANNA.
Статистическая обработка выполнялась с помощью вариационного анализа данных в программе Excel; повторность в опытах четырехкратная.
Результаты исследования и их обсуждение
Данные табл. 1 отражают динамику численности железо- и марганец-окисляющих микроорганизмов, участвующих в деструкции кремнийсодержащих материалов.
Главной выявленной особенностью данных систем явилось двухпиковое увеличение численности клеток, которое приходилось в основном на 5-ый день на варианте с диатомитом (до 130,0×106 и 152,9×106 клеток/мл) и цеолитом (до 374,3×106 и 190,0×106 клеток/мл), а также на 7-ой день на варианте с бентонитовой глиной (до 157,0×106 и 210,4×106 клеток/мл). Второй пик численности приходился, как правило, на 15–20-ый дни культивирования.
На рис. 1 и 2 представлена 30-дневная динамика численности живых клеток накопительной природной культуры соответственно литотрофных и органотрофных фосфатредуцирующих бактерий дерново-подзолистой почвы в системе «порода – культура».
Таблица 1
Динамика численности железо- и марганец-окисляющих бактерий в системе «порода – культура» при деградации высококремнистых пород
|
Численность, ×106 клеток/мл |
День учета жизнеспособности культуры |
|||||||||
|
1 |
3 |
5 |
7 |
10 |
12 |
15 |
20 |
25 |
30 |
|
|
Fe-окисляющие бактерии (АБ) |
||||||||||
|
Диатомит |
22,1 |
54,9 |
130,0 |
86,6 |
63,3 |
92,4 |
180,1 |
240,6 |
92,5 |
44,4 |
|
Цеолит |
48,0 |
120,6 |
374,3 |
200,7 |
28,0 |
151,4 |
184,6 |
48,2 |
16,8 |
1,2 |
|
Бентонит |
36,2 |
66,8 |
100,0 |
157,0 |
63,6 |
84,9 |
101,2 |
64,9 |
36,1 |
12,5 |
|
Mn-окисляющие бактерии (АК) |
||||||||||
|
Диатомит |
12,4 |
32,9 |
152,9 |
73,3 |
91,4 |
214,7 |
604,8 |
96,8 |
60,7 |
32,0 |
|
Цеолит |
62,1 |
80,4 |
190,0 |
154,7 |
120,0 |
361,8 |
456,4 |
150,0 |
96,2 |
60,0 |
|
Бентонит |
22,7 |
71,3 |
170,0 |
210,4 |
110,0 |
90,1 |
128,8 |
325,9 |
148,2 |
84,9 |
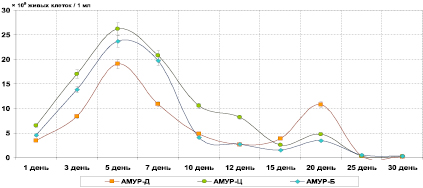
Рис. 1. Численность живых клеток культуры литотрофных (АМУР) фосфатредуцирующих бактерий при биохимической деградации кремнийсодержащих материалов
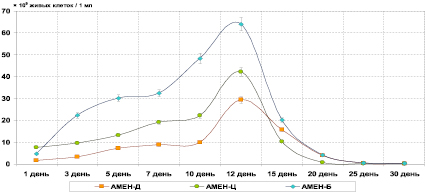
Рис. 2. Численность живых клеток культуры органотрофных (АМЕН) фосфатредуцирующих бактерий при биохимической деградации кремнийсодержащих материалов
Было установлено, что при бактериальной деградации диатомовой породы численность фосфатредуцирующих литотрофных микроорганизмов имела два пика – на 5-ый и 20-ый день культивирования, где составила соответственно 19,14×109 клеток/1 мл и 10,84×109 клеток/1 мл. Между указанными днями учета имелся достаточно значительный спад числа живых клеток бактерий (снижение на 12-ый день до 2,69×109 клеток/1 мл). Численность фосфатредуцирующих органотрофных микроорганизмов, напротив, возрастала достаточно степенно к 10-му дню экспозиции эксперимента (до 9,85×109 клеток/1 мл), а затем давала один существенный подъем жизнеспособности – к 12-му дню до 29,34×109 клеток/1 мл.
В системе «цеолит – культура» наблюдалась аналогичная двухпиковая картина увеличения числа живых клеток у бактерий фосфатредуцентов-литотрофов: повышение численности на 2-ый день до 26,19×109 клеток/1 мл и на 20-ый день до 4,76×109 клеток/1 мл. В опыте с изучением численности фосфатредуцентов-органотрофов на всю экспозицию пришелся один пик максимального числа клеток и, подобно опыту с диатомитом Инзенского месторождения, пришедшийся на 12-ый день – до 42,18×109 клеток/1 мл. В последующие дни опыта жизнеспособность культуры на АМЕН шла на резкий спад – так, на 15-ый день она составила 10,28×109 клеток/1 мл.
В эксперименте с бентонитовой глиной аналогичным образом выстроилась динамика численности живых клеток обеих культур бактерий. В отношении фосфатредуцирующих органотрофов (АМЕН) количество клеток возрастало плавно до 7-го дня (32,46×109 клеток/1 мл) с последующим резким увеличением, пришедшимся на 12-ый день и составившим 64,10×109 клеток/1 мл.
В отношении фосфатредуцирующих органотрофов (АМУР) число живых клеток сильнее возрастало к 5-му дню (23,68×109 клеток/1 мл), а также давало незначительный подъем показателя на 20-ый день – до 3,44×109 клеток/1 мл. На 12-ый и 15-ый дни культивирования пришлась наименьшая численность данных бактерий (2,76 и 1,53×109 клеток/1 мл).
Также необходимо отметить общую для всех систем «порода – культура» особенность повышения численности органотрофных бактерий (АМЕН) после пика численности литотрофных фосфатредуцентов (АМЕН). Вполне вероятно, что данная зависимость могла быть связана с первичным отмиранием литотрофных клеток в культуральной жидкости накопительной культуры и высвобождением органических веществ цитоплазмы в раствор, которые могут являться субстратом для питания органотрофов, что, в свою очередь, и могло приводить к росту жизнеспособности фосфатредуцентов, растущих на агаре Менкиной.
На рис. 3 показана 30-дневная динамика свободной кислотности системы «порода – культура» в зависимости от изучаемого кремнийсодержащего материала и вида фосфатредуцирующих бактерий. Прежде всего, нужно отметить, что вариант с бентонитовой глиной в случае всех действующих на нее микроорганизмов в начале экспозиции эксперимента характеризовался наибольшим значением рН в сравнении с вариантом с диатомовой и цеолитовой породой, что могло быть связано с исходными характеристиками самого бентонита.
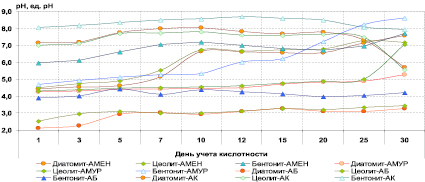
Рис. 3. Динамика свободной кислотности накопительной почвенной культуры фосфатредуцирующих, железо- и марганец-окисляющих бактерий при биохимической деградации высококремнистых пород
По данным рис. 3 видно, что пики максимального увеличения рН систем приходились, как правило, на 10-й день экспозиции, а также на ее конец (30-й день), что, очевидно, могло быть связано как с колебанием численности живых клеток и, как следствие, с метаболизмом в систему продуктов жизнедеятельности, в том числе экзоферментов, полисахаридов и органических кислот.
Также здесь прослеживается то, что кислотность всех бактериальных систем как фосфатредуцентов, так и металлоокислителей стремилась в сторону нейтрального и слабощелочного диапазонов, что, очевидно, связано с растворением Ca- и Mg-содержащих микроструктур исследуемых материалов. В связи с этим можно предположить, что содержание растворимых соединений фосфора, кремния, кальция, магния, а высвобождение микроколичеств марганца и железа в системах «порода – культура» может быть сопряжено не только с прямым действием метаболитов на вещество пород, но в том числе и с образованием нерастворимых оснований, а также фосфатов и карбонатов металлов, последние из которых присутствуют как в составе питательных сред, так и в веществе изучаемых материалов.
В табл. 2 показана динамика общей минерализации культуральной жидкости в системах всех изученных бактериальных комплексов при деградации кремниевых пород.
Исходя из данных таблицы следует, что деятельность Fe-редуцирующих бактерий (АБ) наиболее биохимически агрессивна по отношению к веществу пород – высвобождение в растворимое состояние элементов достигает 480–490 мг/мл. Наименьшая биохимическая активность прослеживается в отношении Mn-редуцентов (АК), где максимальное содержание растворимых форм химических элементов в культуре не превышает и 50–90 мг/мл. Также нужно сказать, что пики наибольших концентраций растворенных соединений в бактериальных суспензиях приходились на 5-ый – 10-ый дни культивирования в зависимости от породы и самой бактериальной культуры.
Заключение
В условиях лабораторных экспозиционных опытов было изучено 30-дневное изменение свободной кислотности культуральной жидкости, а также ее общей минерализации и жизнеспособности в виде численности живых микробных клеток железо-, марганец-, литотрофных и органотрофных фосфатредуцирующих бактерий, происходящее при биохимической деградации природных высококремнистых материалов.
Таблица 2
Изменение общей минерализации культуральной жидкости железо-, марганец- и фосфатредуцирующих бактерий в системе «порода – культура» при деградации высококремнистых пород
|
Общая минерализация, мг/мл |
День учета минерализации |
|||||||||
|
1 |
3 |
5 |
7 |
10 |
12 |
15 |
20 |
25 |
30 |
|
|
Fe-окисляющие бактерии (АБ) |
||||||||||
|
Диатомит |
152 |
212 |
488 |
463 |
423 |
381 |
315 |
301 |
265 |
210 |
|
Цеолит |
156 |
320 |
479 |
452 |
409 |
397 |
348 |
335 |
250 |
200 |
|
Бентонит |
214 |
322 |
485 |
491 |
421 |
388 |
336 |
315 |
263 |
212 |
|
Mn-окисляющие бактерии (АК) |
||||||||||
|
Диатомит |
15 |
33 |
49 |
52 |
49 |
42 |
39 |
35 |
29 |
20 |
|
Цеолит |
30 |
40 |
48 |
46 |
43 |
41 |
39 |
34 |
30 |
28 |
|
Бентонит |
68 |
71 |
86 |
90 |
91 |
83 |
76 |
65 |
47 |
35 |
|
Литотрофные фосфатредуценты (АМУР) |
||||||||||
|
Диатомит |
149 |
217 |
270 |
252 |
231 |
211 |
186 |
141 |
138 |
124 |
|
Цеолит |
110 |
250 |
315 |
396 |
288 |
262 |
211 |
183 |
91 |
86 |
|
Бентонит |
83 |
142 |
264 |
323 |
382 |
290 |
271 |
267 |
119 |
79 |
|
Органотрофные фосфатредуценты (АМЕН) |
||||||||||
|
Диатомит |
110 |
161 |
227 |
248 |
184 |
176 |
148 |
136 |
98 |
64 |
|
Цеолит |
92 |
120 |
205 |
199 |
179 |
155 |
121 |
116 |
83 |
30 |
|
Бентонит |
71 |
121 |
243 |
251 |
222 |
201 |
183 |
137 |
106 |
74 |
Было установлено, что численность фосфатредуцентов-литотрофов имела наибольшее увеличение при деградации цеолитовой породы, оказалась на среднем уровне при деструкции бентонитовой глины и была минимальной при минерализации диатомита. Фосфатредуцирующие органотрофные бактерии давали максимальную численность на варианте с бентонитом, среднюю – на варианте с цеолитом и минимальную – на варианте с диатомовой породой. Было выявлено, что экспозиция эксперимента с бактериями литотрофного типа питания (в том числе с Fe- и Mn-окисляющей культурами) давала двухпиковую жизнеспособность клеток вне зависимости от изучаемой породы, в то время как в опытах с бактериями органотрофного типа питания один пик численности стабильно приходился на 12–15-ый дни эксперимента.
Максимальные концентрации растворенных соединений (общая минерализация культуральной жидкости) в бактериальных суспензиях приходились на 5-ый – 10-ый дни культивирования в зависимости от породы и самой бактериальной культуры и достигала 480–490 мг/мл по культуре железо-редуцентов, 50–90 мг/мл – по культуре марганец-редуцентов, 252–396 мг/мл – по культуре фосфатредуцентов-литотрофов и 205–251 мг/мл по культуре фосфатредуцентов-органотрофов. Также установлено, что кислотность всех бактериальных систем, проявившая пики на 10-й и 30-й дни экспозиции экспериментов, стремилась в сторону нейтрального и слабощелочного диапазонов, что, очевидно, связано с растворением Ca- и Mg-содержащих микроструктур исследуемых кремнийсодержащих материалов.
Библиографическая ссылка
Козлов А.В., Уромова И.П. ИЗМЕНЕНИЕ ПАРАМЕТРОВ МИКРОБИОЛОГИЧЕСКОЙ СИСТЕМЫ ЖЕЛЕЗО-, МАРГАНЕЦ- И ФОСФАТРЕДУЦИРУЮЩИХ БАКТЕРИЙ ПРИ ДЕСТРУКЦИИ ДИАТОМИТА, ЦЕОЛИТА И БЕНТОНИТОВОЙ ГЛИНЫ // Международный журнал прикладных и фундаментальных исследований. 2017. № 8-1. С. 110-115;URL: https://applied-research.ru/en/article/view?id=11770 (дата обращения: 29.06.2026).