
Scientific journal
International Journal of Applied and fundamental research
ISSN 1996-3955
ИФ РИНЦ = 0,556
ABSORPTION OF UV BY BARLEY AT DIFFERENT STAGES OF ORGANOGENESIS

В настоящее время при изучении эффектов ультрафиолетового излучения на биологические объекты используется концепция биологически эффективного УФ-излучения. Данная концепция заключается в оценке относительного вклада в наблюдаемый биологический эффект ультрафиолета произвольной длины волны по отношению к квантам УФИ с определенной длиной волны, вызывающим биологический эффект по величине, принятой за единицу. Спектры биологического действия УФ-В-излучения на морфологические и ростовые характеристики высших растений были разработаны [1] и впоследствии дополнены для УФ-А-излучения [2].
Биологически эффективные дозы, полученные для заданного биологического эффекта, позволяют сравнивать степень радиационного воздействия по величине именно этого биологического эффекта, то есть определять зависимости «доза – эффект» как от искусственных источников ультрафиолетового излучения, так и от солнечного излучения. Но в то же время концепция биологически эффективной дозы имеет ряд ограничений, обусловленных различными механизмами действия ультрафиолета с разной частотой. Поэтому необходимо развивать в дополнение к концепции биологически эффективной дозы также и концепцию поглощенной дозы в отношении неионизирующего излучения.
По мнению одних авторов [3], поглощение лучистой энергии листьями растений происходит в соответствии с законом поглощения Ламберта – Бэра, тогда как другие авторы [4–5] полагают, что этот закон не приложим к рассеивающим биологическим объектам, особенно таким, как листья растений.
В связи с высокой степенью гетерогенности листьев растений оптические и фотометрические закономерности, характеризующие их, безусловно, отличаются от закономерностей, принятых в оптике и спектроскопии прозрачных сред [5]. Поэтому представляет интерес проследить, существует ли закономерная зависимость между биологическим эффектом и величиной коэффициента поглощения лучистой энергии [5].
Однако геометрическая сложность растений, как объекта изучения, не позволяет определить конкретную величину поглощенной энергии, тем более что коэффициент поглощения сильно зависит от спектра излучения.
Цель работы состоит в оценке доли поглощенного ультрафиолетового излучения посевами ячменя на разной стадии онтогенеза.
Материалы и методы исследования
Сущность метода состоит в измерении потока излучения через замкнутую поверхность (в нашем случае – параллелепипед), внутри которой помещен объект изучения, и сравнение с тем же потоком, но без объекта. Объектом изучения были посевы ячменя сорта Зазерский в вегетационном сосуде (диаметром 20 см на уровне почвы) на разной стадии вегетации, плотность посевов составляла 14–20 растений на сосуд, или 350–480 растений на метр квадратный, в одном случае, для сравнения, использовали разреженный посев – 7 растений на сосуд, или 170 растений на метр квадратный. Для фиксации радиометра в постоянных точках измерения использовали параллелепипед, изготовленный из металлической сетки размером ячейки 45х45 мм, что обусловлено конструктивными особенностями радиометра. Вегетационный сосуд с посевами ячменя размещали таким образом, чтобы измерения на нижней грани параллелепипеда приходились на уровне почвы. Для фоновых измерений таким же образом размещали вегетационный сосуд с почвой без растений. Источником УФ-излучения являлась лампа ДРТ-240, размещенная в 30 см над верхней поверхностью параллелепипеда по центру, измерения проводили в темной комнате при стабильном режиме работы лампы.
Измерение потока излучения проводили УФ-радиометром ТКА-АВС, предназначенном для измерения энергетической УФ-облученности в трех диапазонах:
– УФ-А (315…400 нм) источниками УФ-излучения за исключением газоразрядных ртутных ламп без люминофоров;
– УФ-В (280…315 нм) источниками УФ-излучения за исключением газоразрядных ртутных ламп с люминофорами типа «А», а также естественных источников излучения;
– УФ-С (200…280 нм) газоразрядными ртутными лампами высокого и низкого давления без люминофоров.
Поскольку в нашем эксперименте источником излучения являлась ртутная лампа высокого давления без люминофора, показания прибора в области УФ-А имели систематическую погрешность. Однако, так как нас интересует только относительная доля поглощенного УФ-излучения, данное обстоятельство не имеет значения.
Угол измерения данным прибором составляет 20–30 °, в зависимости от диапазона, поэтому в некотором приближении можно считать показания измерения равными потоку излучения, перпендикулярному плоскости измерения.
Измерения проводили по всем ячейкам параллелепипеда 7х7х11. Для дальнейших расчетов использовали только те ячейки, разница в которых при измерении с объектом составляла более 5 % от фоновых измерений.
Поглощенную мощность излучения рассчитывали как разницу между не прошедшей через посевы и отраженной мощностью излучений:
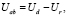 (1)
(1)
где Ud – не прошедший через посевы поток УФ-излучения, интегрированный по площади тех ячеек с объектом, где поток уменьшался, разница с фоновыми измерениями, мВт;
Ur – отраженный посевами поток УФ-излучения, интегрированный по площади тех ячеек с объектом, где поток увеличивался, разница с фоновыми измерениями, мВт.
Поскольку источник освещения располагался на небольшом расстоянии от объекта, средний поток излучения на посевы рассчитывали по формуле
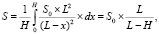 (2)
(2)
где S0 – поток излучения на уровне почвы, Вт/м2;
L – расстояние от почвы до источника излучения, м;
Н – средняя высота растений в посевах, м.
Для сравнения различных измерений и эффективности поглощения УФ разного диапазона полученные результаты нормировали на одно растение и на поток излучения 1 Вт/м2 на уровне почвы.
Достоверность различий средних значений проводили t-критерием, в тексте и таблице представлены средние значения с ошибкой.
Результаты исследования и их обсуждение
На рис. 1 представлен пример фактических измерений потока УФ-излучения через поверхность параллелепипеда при фоновом измерении и с объектом. Как и следовало ожидать, максимальная часть потока проходит через нижнюю грань, на боковых гранях наблюдается сверху вниз снижение тангенциальной доли излучения от источника, а на верхней грани регистрируется незначительный поток отраженного излучения от внутренних элементов установки.
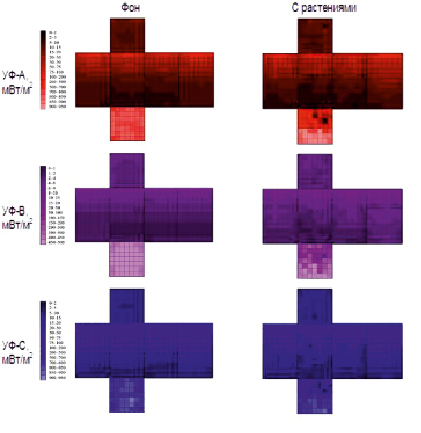
Рис. 1. Развертка поверхности измерения потока УФ-излучения для посевов ячменя на стадии колошения
Регистрация изменения потока при измерении с объектом (посевы ячменя), хорошо видимая на рис. 1, позволила рассчитать мощность поглощенного и отраженного УФ-излучения. Полученные данные, а также морфологические показатели посевов ячменя представлены в табл. 1–2. Следует отметить, что при вегетационных экспериментах в условиях теплицы развитие растений проходит заметно медленнее, чем должно быть в полевых условиях, к тому же образование боковых стеблей и их рост сильно замедлены.
Таблица 1
Показатели растений ячменя в посевах
|
№ измерения |
Возраст растений, сутки |
Количество растений на сосуд |
Этап органогенеза по Куперману |
Высота растений, см |
Общая масса листьев, г |
Надземная фитомасса, г |
Общая площадь листьев, см2 |
|
1 |
20 |
19 |
II |
23,4 ± 1,5 |
5,32 |
7,98 |
76 ± 6 |
|
2 |
29 |
20 |
II |
31,6 ± 1,4 |
5,8 |
9,4 |
98 ± 7 |
|
3 |
35 |
7 |
III |
49,1 ± 1,5 |
11,2 |
18,2 |
89 ± 8 |
|
4 |
58 |
18 |
V–VI |
48,1 ± 1,3 |
13,68 |
24,84 |
140 ± 12 |
|
5 |
86 |
20 |
VIII |
60,3 ± 3,0 |
16,8 |
47,5 |
240 ± 20 |
|
6 |
100 |
14 |
X |
64,4 ± 2,0 |
29,4 |
71,1 |
239 ± 14 |
Таблица 2
Поглощенная и отраженная мощность УФ-излучения в посевах ячменя
|
№ измерения |
Поток на посев, Вт/м2 |
Поглощенная мощность УФ, мВт по диапазонам |
Отраженная мощность УФ, мВт по диапазонам |
||||||
|
А |
В |
С |
А |
В |
С |
А |
В |
С |
|
|
1 |
1,4 |
0,7 |
1,2 |
4,4 |
0,4 |
7,0 |
0,1 |
0,04 |
0,4 |
|
2 |
1,6 |
0,8 |
1,4 |
14,2 |
6,9 |
11,5 |
0,3 |
0,1 |
0,7 |
|
3 |
2,6 |
1,3 |
2,3 |
20,3 |
12,5 |
24,2 |
0,6 |
0,3 |
1,5 |
|
4 |
2,6 |
1,4 |
2,3 |
43,1 |
20,2 |
36,6 |
0,6 |
0,3 |
1,5 |
|
5 |
4,7 |
2,5 |
4,1 |
78,5 |
39,1 |
66,1 |
2,0 |
0,9 |
1,0 |
|
6 |
5,9 |
3,0 |
5,0 |
57,1 |
30,7 |
47,8 |
1,9 |
1,3 |
2,1 |
Нормирование полученных данных на одно растение в посеве и на поток УФ-излучения в 1 Вт/м2 позволяет сравнить поглощение и отражение ультрафиолета различных диапазонов ячменем на разных этапах органогенеза (табл. 3). Ранговый дисперсионный анализ данных табл. 3 показывает отсутствие различий как в поглощении (p < 0,40), так и в отражении (p < 0,22) УФ разных диапазонов. В то же время различия между измерениями высоко достоверны (p < 0,01).
Таблица 3
Поглощенная и отраженная мощность УФ-излучения отдельным растением ячменя при потоке 1 Вт/м2
|
№ измерения |
Поглощенная мощность УФ, мВт по диапазонам |
Отраженная мощность УФ, мВт по диапазонам |
||||||
|
А |
В |
С |
Среднее |
А |
В |
С |
Среднее |
|
|
1 |
0,17 |
0,03 |
0,31 |
0,17 ± 0,08 |
0,004 |
0,003 |
0,018 |
0,008 ± 0,005 |
|
2 |
0,44 |
0,41 |
0,41 |
0,42 ± 0,01 |
0,009 |
0,006 |
0,025 |
0,013 ± 0,006 |
|
3 |
1,11 |
1,35 |
1,52 |
1,33 ± 0,12 |
0,033 |
0,032 |
0,094 |
0,053 ± 0,021 |
|
4 |
0,92 |
0,82 |
0,89 |
0,88 ± 0,03 |
0,013 |
0,012 |
0,036 |
0,020 ± 0,008 |
|
5 |
0,84 |
0,80 |
0,81 |
0,82 ± 0,01 |
0,021 |
0,018 |
0,012 |
0,017 ± 0,003 |
|
6 |
0,69 |
0,72 |
0,68 |
0,70 ± 0,01 |
0,023 |
0,031 |
0,030 |
0,028 ± 0,002 |
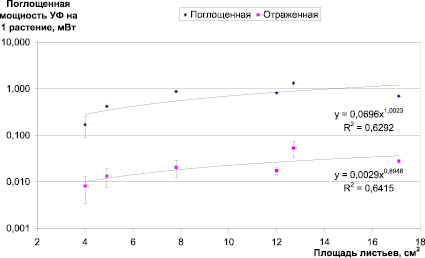
Рис. 2. Зависимость поглощенной и отраженной мощности УФ-излучения ячменем от площади листьев растения
Различия в величине поглощенного УФ на разных этапах органогенеза растений обусловлены в первую очередь изменениями в величине ассимилирующей поверхности, площади листьев и их расположением (рис. 2).
Если рассматривать поглощенный и отраженный поток ультрафиолета с учетом площади листовой поверхности, то можно убедиться, что на определенном этапе органогенеза растение поглощает практически все, падающее на него УФ-излучение (рис. 3).
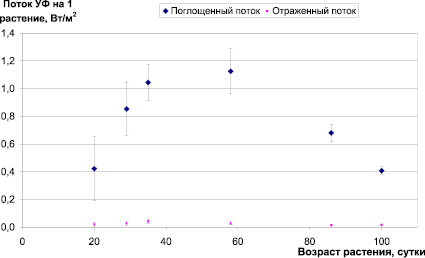
Рис. 3. Изменение удельного поглощенного и отраженного ультрафиолета от возраста растений ячменя при потоке УФ-излучения на уровне почвы 1 Вт/м2
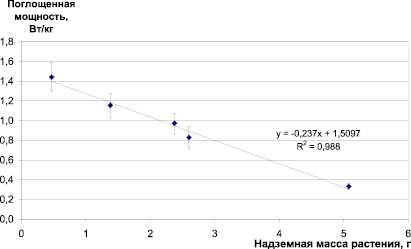
Рис. 4. Удельная поглощенная мощность УФ-излучения (Вт/кг) при плотности потока 1 Вт/м2 в зависимости от надземной биомассы растений ячменя
Как видно из данных, представленных на рис. 3, на III–VI этапах органогенеза растение поглощает практически весь падающий на него поток (1 Вт/м2), отраженный поток не зависит достоверно от этапа органогенеза и составляет в среднем всего 2,4 ± 0,4 % от потока, падающего на растение. Низкая удельная доля поглощенного УФ-излучения на раннем этапе органогенеза обусловлена, по-видимому, вертикальным расположением листьев растения, что снижает их проективное покрытие по сравнению с площадью листьев. На поздних этапах органогенеза снижение удельной доли поглощенного ультрафиолета обусловлено частичным затенением листьев в посевах.
Подтверждением этого предположения является наличие большого количества защитных пигментов в клетках листьев ячменя. Подобного мнения придерживаются и другие авторы [5]. Ряд авторов сообщали, что растения поглощают УФ-радиацию очень эффективно (свыше 90 %), и что в сущности ее не пропускают [6]. Следовательно, обычно отражается менее чем 10 % УФ-радиации. Интересно заметить, что в случае пустынных сочных растений, таких как Agave americana и Opuntia leavis, отражение может достигать 10–20 %, в то время как у хвойных растений отражение может быть только 2–3 % [6].
В настоящее время в радиобиологии неионизирующих излучений основным дозовым критерием является экспозиционная доза (Дж/м2), однако радиобиологические эффекты обусловлены не экспозиционной, а поглощенной дозой излучения (Дж/кг). Учитывая, что поглощение излучения у растений ячменя происходит в основном листьями, как имеющими наибольшую ассимилирующую поверхность, можно рассчитать удельную поглощенную мощность при потоке 1 Вт/м2. При этом, если рассматривать поглощенную мощность УФ-излучения в листьями ячменя в зависимости от общей надземной биомассы растений (рис. 4), можно отметить быстрое снижение удельной поглощенной дозы по мере роста растения.
Таким образом, оценивая радиобиологические эффекты от действия УФ-радиации на ячмень, необходимо учитывать различия в поглощении ультрафиолета на разных этапах органогенеза растений.
Выводы
1. Различные диапазоны УФ-излучения практически одинаково поглощаются и отражаются растениями. При этом доля отраженного ультрафиолета для ячменя составляет 2,4 %.
2. Зависимость поглощенной доли ультрафиолета от общей площади листьев носит линейный характер и обусловлена увеличением проективного покрытия по мере роста растений и увеличения количества листьев. На ранних и поздних этапах органогенеза растений ячменя поглощение ультрафиолетового излучения значительно снижено, по сравнению с III–VI этапами органогенеза, что обусловлено различным пространственным расположением листьев в посевах.
3. Для сравнения радиобиологических эффектов действия острого УФ-облучения на растения, находящихся на разных этапах органогенеза, необходимо учитывать не только экспозиционную, но и поглощенную дозу.
Библиографическая ссылка
Цыгвинцев П.Н. ПОГЛОЩЕНИЕ УФ ЯЧМЕНЕМ НА РАЗНЫХ ЭТАПАХ ОРГАНОГЕНЕЗА // Международный журнал прикладных и фундаментальных исследований. 2017. № 10-2. С. 283-288;URL: https://applied-research.ru/en/article/view?id=11905 (дата обращения: 09.07.2026).
DOI: https://doi.org/10.17513/mjpfi.11905