
Scientific journal
International Journal of Applied and fundamental research
ISSN 1996-3955
ИФ РИНЦ = 0,556
DYNAMICS OF INDEXES OF A VARIETY OF COMMUNITIES OF BATS (CHIROPTERA, VESPERTILIONIDAE) ON HIBERNATION IN ARTIFICIAL UNDERGROUND

Рукокрылые (Сhiroptera) представлены в фауне России семейством Гладконосые (Vespertilionidae). На Южном Урале обитает 16 видов летучих мышей этого семейства [1–3], из них 7 видов относятся к группе перелетных, а 9 – к группе оседлых, зимующих в местах летнего обитания. За период активной индустриализации ХХ века на Урале кроме естественных карстовых полостей (пещер), используемых рукокрылыми для зимовки, появились искусственные техногенные подземелья (шахты и штольни) – результат добычи горных пород и минералов. Изучение заселения рукокрылыми техногенных подземелий и изменений биоразнообразия на зимовках позволяет выявить успешность их функционирования в антропогенно-трансформированной среде.
В качестве модельного подземелья исследовали штольни пос. Слюдорудник (Челябинская обл.). Штольни образованы в период 1930–1965 гг. в результате подземных разработок. По своим микроклиматическим условиям (температура от –8 до +7 °С, влажность 50–99 %) и линейным характеристикам (общая протяженность ходов около 4 км) штольни пригодны для зимовок рукокрылых. Целью работы было выявление фаунистического состава и оценка многолетних изменений биоразнообразия рукокрылых на зимовке в техногенном подземелье.
Материалы и методы исследования
Материалом работы послужили данные зимних учетов летучих мышей, полученные в 2007–2008 гг. и 2010–2017 гг. в штольнях пос. Слюдорудник. Летучих мышей учитывали визуально. Видовую принадлежность определяли при осмотре по внешним характерным признакам [4, 5], иногда для уточнения вида животных извлекали из укрытий. Ночниц, которых точно определить не могли, отмечали как Myotis sp. Для оценки количественного соотношения видов в сообществе использовали показатель относительного обилия (D), определяемый как отношение числа особей данного вида (N) к общему числу особей всех видов, отмеченных в одной выборке (S): D = N / S.
Биоразнообразие сообществ рукокрылых оценивали с помощью информационного индекса Шеннона (H’), характеризующего представленность видов в сообществе по численности [6–8]:
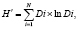
где Di – доля особей i-го вида (относительное обилие).
В дополнение к индексу Шеннона вычисляли показатель выравненности сообщества по Пиелу (Е): чем более равномерно представлены в сообществе составляющие его виды, тем ближе его значение к единице:
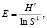
где S – общее число видов в выборке.
Расчеты осуществляли с использованием пакета Microsoft Excel и Statistica 6.1 for Windows.
Результаты исследования и их обсуждение
В ходе исследований было учтено 709 летучих мышей 6 видов: северный кожанок Eptesicus nilssonii, бурый ушан Plecotus auritus, ночница водяная Myotis daubentonii, ночница Брандта M. brandtii, ночница прудовая M. dasycneme и ночница усатая M. mystacinus (табл. 1).
Таблица 1
Видовой состав, численность (N) и относительное обилие (D) рукокрылых на зимовках в 2006/2007–2016/2017 гг. в штольнях пос. Слюдорудник
|
Зимовки, годы |
E. nilssonii |
Pl. auritus |
M. daubentonii |
M. brandtii |
M. mystacinus |
M. dasycneme |
Myotis sp. |
S |
|||||||
|
N |
D |
N |
D |
N |
D |
N |
D |
N |
D |
N |
D |
N |
D |
||
|
2006/2007 |
28 |
0,24 |
33 |
0,28 |
15 |
0,13 |
34 |
0,29 |
1 |
0,01 |
0 |
0 |
5 |
0,04 |
116 |
|
2007/2008 |
42 |
0,32 |
20 |
0,15 |
13 |
0,10 |
40 |
0,30 |
0 |
0 |
6 |
0,05 |
11 |
0,08 |
132 |
|
2010/2011 |
12 |
0,12 |
2 |
0,02 |
14 |
0,14 |
65 |
0,66 |
0 |
0 |
6 |
0,06 |
1 |
0,01 |
99 |
|
2012/2013 |
24 |
0,20 |
39 |
0,32 |
21 |
0,17 |
34 |
0,28 |
0 |
0 |
3 |
0,02 |
1 |
0,01 |
122 |
|
2014/2015 |
47 |
0,34 |
12 |
0,09 |
24 |
0,18 |
50 |
0,36 |
0 |
0 |
4 |
0,03 |
1 |
0,01 |
138 |
|
2016/2017 |
23 |
0,23 |
14 |
0,14 |
21 |
0,21 |
33 |
0,33 |
2 |
0,02 |
7 |
0,07 |
2 |
0,02 |
102 |
|
Всего |
176 |
120 |
108 |
256 |
3 |
26 |
20 |
709 |
|||||||
Ежегодные количества учтенных животных в исследовании близки по объему и достаточны (около 100 экз.), что репрезентативно для расчетов информационных индексов [8, 9]. Для проверки на нормальность распределения количественных данных рассчитали математическое ожидание (М), среднее квадратичное отклонение (s), медиану, моду изменяющейся численности каждого вида летучих мышей (табл. 2).
Таблица 2
Данные описательной статистики численности видов рукокрылых на зимовках 2006/2007–20016/2017 гг. в штольнях пос. Слюдорудник
|
Виды |
Число наблюдений |
М ± s |
Min |
Max |
Медиана |
Мода |
|
Myotis daubentonii |
6 |
18,00 ± 4,56 |
13 |
24 |
18 |
21 |
|
Myotis dasycneme |
6 |
4,33 ± 2,58 |
0 |
7 |
5 |
6 |
|
Myotis brandtii |
6 |
42,67 ± 12,68 |
33 |
65 |
37 |
34 |
|
Myotis mystacinus |
6 |
0,50 ± 0,84 |
0 |
2 |
0 |
0 |
|
Eptesicus nilssonii |
6 |
29,33 ± 12,99 |
12 |
47 |
26 |
множ. |
|
Plecotus auritus |
6 |
20,00 ± 13,81 |
2 |
39 |
17 |
множ. |
Полученные данные по численности видов рукокрылых в штольнях на зимовке не симметричны относительно своего среднего значения M: математическое ожидание, мода и медиана не совпадают. Но большинство значений попадает в интервал среднего квадратичного отклонения (M ± s). Превышение величины s над M отмечено у редкого вида M. mystacinus. Медианные значения численности доминирующих видов рукокрылых – E. nilssonii, Pl. auritus, M. brandtii – близки были в период исследований к нижнему квартилю. Попадание медианы внутрь диапазона квартильного размаха численности выполняется для всех видов летучих мышей за исключением M. mystacinus (рис. 1).
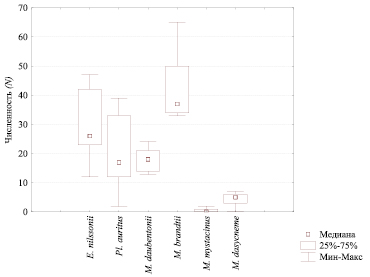
Рис. 1. Соотношение медианы и квартильного размаха численности каждого вида летучих мышей, учтенных на зимовках в период 2006/2007–2016/2017 гг. в штольнях пос. Слюдорудник
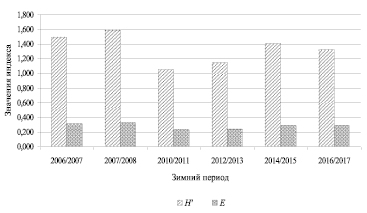
Рис. 2. Многолетняя динамика индекса биоразнообразия Шеннона (H′) и показателя выровненности Пиелу (Е) сообществ рукокрылых на зимовках в штольнях пос. Слюдорудник
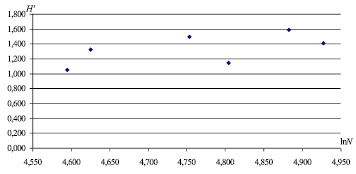
Рис. 3. Зависимость между численностью (ln N) и значениями индекса разнообразия Шеннона (H′)
Из анализа численности следует, что к полученным данным можно применять расчетные методы вычисления индексов биоразнообразия, используя в расчетных формулах относительное обилие и «нормализуя» логарифмированием (ln Di). Отсутствие нормальности распределения численности может свидетельствовать о нахождении сообщества рукокрылых на зимовке под воздействием стресса факторов среды [8, 9]. Из данного заключения можно сделать вывод: рассчитанные индексы биоразнообразия (H’) сообществ рукокрылых на зимовках варьируют в пределах 1,1–1,6, их изменчивость в межгодовой динамике представлена на рис. 2.
В различных сообществах значения индекса лежат в пределах от 1,5 до 3,5, редко превышая 4,5 [8, 9], полученные нами значения находятся у нижних пределов. Невысокие значения индексов биоразнообразия могут указывать на нестабильное состояние сообщества рукокрылых. Известно, что индекс Шеннона суммирует большое количество информации о численности, видовом составе и сильно зависит от числа редких видов, характеризуя видовое богатство: снижение индекса мы наблюдали при выпадение редких видов из учетов (M. mystacinus). Исследуемое сообщество рукокрылых является маловидовым (S = 6), а в маловидовых сообществах (≤ 30) на величину индекса преимущественно влияет количество видов, а не их представленность в сообществе. Невысокое значение индекса Шеннона указывает на неустойчивое состояние сообщества рукокрылых [8, 9]. Мы связываем неустойчивость сообщества, характеризуемую низким индексом биоразнообразия, с недавним заселением штолен рукокрылыми с 1970 годов. Многолетняя динамика показывает снижение индексов биоразнообразия на зимовке в 2010/2011 гг. (рис. 2), что могло происходить вследствие дополнительного воздействия внешних неблагоприятных факторов, приводящего к увеличению стрессового состояния сообществ рукокрылых. Зависимость величины индекса разнообразия Шеннона от численности сообщества (ln N) представлена на рис. 3.
Из рисунка видно, что зависимость величины индекса Шеннона от численности сообщества не наблюдается, что свидетельствует о репрезентативности данных учетов и достоверности многолетних исследований индекса Шеннона.
Использование индекса Пиелу позволило оценить изменчивость выравненности видов в сообществах зимовки по их относительному обилию. Значения индекса Пиелу варьировали в узких пределах 0,239–0,326, характеризуя постоянно низкую выровненность видов в сообществе, наблюдается выраженное преобладание в сообществе одних видов над другими. Наибольшие величины этого индекса отмечены на зимовке 2007/2008 гг. Прослеживается рост выровненности сообществ (Е) на зимовках, с сезона 2010/2011 по 2016/2017 гг. Похожие тенденции отмечены для населения рукокрылых, зимующих в штольнях Самарской Луки в Поволжье, заселяемых во второй половине XX века [10, 11]: к концу века наблюдался максимум видового разнообразия, выровненности сообществ. Исследуемые нами штольни начали заселяться позже, и пока мы наблюдаем колебания биоразнообразия. Низкое значение индекса Пиелу, свидетельствующее о сильном доминировании одних видов над другими, может характеризовать больший уровень экологической адаптации некоторых видов (E. nilssonii, Pl. auritus, M. brandtii) к условиям техногенных подземелий.
Заключение
Таким образом, в ходе исследования на зимовке в техногенном подземелье было учтено 709 особей 6 видов летучих мышей. Три вида являются обычными и многочисленными на зимовках и входят в доминирующий комплекс рукокрылых подземелья – северный кожанок E. nilssonii, бурый ушан Pl. auritus, ночница Брандта M. brandtii. Два вида являются редкими и встречаются с малой численностью на зимовке: ночницы прудовая M. dasycneme и усатая M. mystacinus. Обнаружено выраженное преобладание одних видов летучих мышей над другими – постоянно низкую выровненность видов в сообществе характеризовали низкие значения индекса разнообразия Пиелу (0,2–0,3). Индекс разнообразия Шеннона имел низкие значения (1,1–1,6), что указывает на нестабильное состояние сообщества рукокрылых: стрессовое состояние вследствие расширения ареала, адаптации к новым антропогенным (техногенным) ландшафтам. Динамика индексов разнообразия показывает его заметное снижение на зимовке 2010/2011 гг., что связано с природно-климатическими факторами – температурой и влажностью, влияющими на промерзание штолен в зимний период.
Библиографическая ссылка
Снитько К.В., Снитько В.П. ДИНАМИКА ИНДЕКСОВ РАЗНООБРАЗИЯ СООБЩЕСТВ РУКОКРЫЛЫХ (CHIROPTERA, VESPERTILIONIDAE) НА ЗИМОВКЕ В ТЕХНОГЕННОМ ПОДЗЕМЕЛЬЕ // Международный журнал прикладных и фундаментальных исследований. 2018. № 1. С. 126-130;URL: https://applied-research.ru/en/article/view?id=12078 (дата обращения: 22.06.2026).
DOI: https://doi.org/10.17513/mjpfi.12078