
Scientific journal
International Journal of Applied and fundamental research
ISSN 1996-3955
ИФ РИНЦ = 0,556
DEVELOPMENT OF MORPHOFUNCTIONAL MECHANISMS OF THE ADAPTATIONS OF SAIGA PREGASTRICS IN ONTOGENESIS TO HABITAT CONDITIONS

Приспособительные механизмы жвачных животных к условиям обитания были и остаются среди приоритетных направлений эволюционно-морфологических исследований. Наиболее существенной формой связи животного организма с окружающей средой, обеспечивающей поддержание энергетического баланса, является связь через пищу. Очевидна роль органов пищеварительной системы животных, обеспечивающих и реализующих эту связь. У дикой фауны в процессе индивидуального и исторического развития приспособительные механизмы оттачивались естественным отбором. Наиболее ценные приспособления среди растительноядных животных приобрели жвачные с обретением сложного многокамерного желудка с главенствующей ролью преджелудочной ферментации [1–3]. Доказано, что желудочно-кишечный тип пищеварения более выражен у диких жвачных, по сравнению с домашними, у которых благодаря вмешательству человека происходит его смягчение и направление в сторону кишечного типа [3, 4]. Исследованию сложного желудка жвачных, процессам пищеварения в них, а также гистофизиологии камер, посвящено множество работ, однако в силу востребованности и доступности материала они касаются домашних жвачных [5–7]. По этой же причине онтогенетические исследования эмбриогенеза камер желудка жвачных, особенно диких видов, единичны [8–10]. К диким видам жвачных копытных, сохранившихся с древнейших времен, благодаря высокой степени адаптивности, относится европейский сайгак – Saiga tatarica Linnaeus. В выживании сайгаков и поддержании энергетического баланса в неблагоприятных аридных условиях, с резкими колебаниями кормовой базы, основная роль принадлежит системе органов пищеварения, в которой главенствуют преджелудки.
Исходя из этого, изучение развития адаптивных механизмов камер желудка сайгаков в течение индивидуального развития, обеспечивающих им выживаемость, является актуальным. Кроме того, за последнее время состояние европейской популяции сайгака, обитающей в Северо-Западном Прикаспии, резко ухудшилось, что придает особую важность их изучению.
Цель исследования: исследование особенностей макро- и микроморфологии преджелудков сайгаков, а также их функциональной активности на протяжении онтогенеза для выяснения развития механизмов адаптации к условиям обитания.
Материалы и методы исследования
Материалом для исследований послужили преджелудки сайгаков: рубец, сетка и книжка от 30 животных: 10 плодов, 5 новорожденных, 5 четырехмесячных и 10 взрослых. Исследования проведены с помощью комплекса гистологических, гистохимических и биохимических методов.
Для гистологического анализа образцы камер желудка сайгаков фиксировали в жидкости Гелли (жидкость Ценкера с формалином) и окрашивали методом Доминичи – Кедровского. Участие в обменных процессах расценивали по наличию, количеству и характеру локализации углеводов, жиров и ферментов: гликогена, гликопротеинов, липазы, сукцинатдегидрогеназы, кислой и щелочной фосфатаз, выявленных классическими методами: Шабадаша, Шубича, Дроздова, Шамахмудова, Гомори, Нахласа, Гроуфорда, Зелигмана [10].
Результаты исследования и их обсуждение
Гистологическими методами выявлено, что у плодов сайгаков в 45 дней (в начале плодного периода развития) стенка преджелудков уже имела дефинитивную форму и состояла их трех оболочек: слизистой, мышечной и серозной (рис. 1). Слизистая оболочка стенки преджелудков образована из многослойного эпителия и соединительной ткани, мышечная – из двух слоев гладких мышечных клеток, серозная – из рыхлой соединительной ткани и тонкого слоя клеток мезотелия. На периферии эпителиального пласта имелись многочисленные экскреторные полости, достигавшие иногда значительных размеров, принимавших форму колб или больших цистерн, открывавшихся наружу, свидетельствуя об участии в обменных функциях через экскрецию (рис. 1).
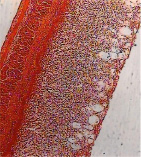
а) б) в)
Рис. 1. Архитектоника слоев стенок преджелудков ранних плодов сайгаков и экскреторные вакуоли эпителиального пласта: а) рубец, б) сетка, в) книжка. Ув. х100
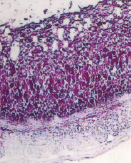
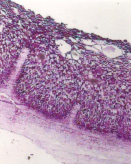
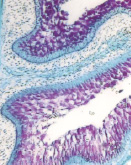
а) б) в)
Рис. 2. Экскреторные полости и гликоген в пузырчатых клетках эпителия преджелудков ранних плодов сайгаков: а) рубец, б) сетка, в) книжка. Ув. х100
Структурные преобразования слизистой оболочки преджелудков плодов сайгаков начинались в разное время – в рубце в 90 дней, сетке – в 60 дней, а в книжке – к началу плодного периода уже имелись листочки первого порядка. В это время происходило прогибание базальной мембраны в сторону поверхности эпителия, образуя соединительнотканные выпячивания – основу будущих морфофункциональных структур преджелудков: сосочков рубца, перегородок сетки, листочков книжки.
После рождения сайгачат происходила постепенная дефинитивная перестройка эмбрионального эпителиального пласта в многослойный слабоороговевающий эпителий, с уменьшением количества вакуолей и образованием гликокаликса, что особенно стимулировалось началом поедания растительного корма. У четырехмесячных сайгачат структура стенок преджелудков полностью имела дефинитивную структуру.
Многослойный эмбриональный эпителий слизистой оболочки преджелудков ранних плодов сайгаков состоял в основном из пузырчатых клеток с небольшим количеством митохондрий и эндоплазматического ретикулума, расположенных на периферии и околоядерной зоне. Верхние края соседних эпителиоцитов соединялись десмосомами, между которыми имелись многочисленные межклеточные щели и микроворсинки.
В позднеплодном периоде (90 дней – рождение) происходило мощное утолщение эпителиального пласта, имевшего до 50 рядов пузырчатых клеток, образование особой складчатости проксимальной поверхности базальных эпителиоцитов, увеличение ширины межклеточных щелей и активное образование цитоплазматических микровыростов. Такое строение эпителия связано не только с увеличением прочности взаимосвязи эпителиоцитов между собой, обеспечивающем защитные свойства, но и с интенсивной двухсторонней транспортной активностью всего эпителиального пласта.
Эпителий слизистой оболочки преджелудков уже в раннеплодный период обладал гликолитической активностью. Об этом свидетельствовало обнаружение по всему пласту гликогена, локализующегося в базальных частях и оболочках эпителиоцитов (рис. 2). Это свидетельствует об участии эпителия преджелудков плодов сайгаков в углеводном обмене, выполняя энергетическую и пластическую функции через расходование на рост и дифференцировку эпителия и участие в формировании гликокаликса.
Результаты биохимического анализа гликогена в преджелудках сайгаков в течение онтогенеза выявили интересную динамику его накопления и расходования (рис. 3). У ранних плодов во всех преджелудках превалировали процессы накопления гликогена над расходованием его на рост и дифференцировку, что приводило к достижению его максимума к 120 дням. По нашему мнению, это является проявлением компенсаторной функции по отношению к печени, гликогендепонирующая функция которой пока низкая и недостаточная для обеспечения обменных функций плода в целом. К рождению, с мощным усилением дифференцировки всех структур слизистых оболочек и повышением гликогендепонирующей функции печени, процессы расходования гликогена в преджелудках начали превалировать над накопительными, что приводило к резкому уменьшению его количества.
После рождения тенденция к уменьшению количества гликогена в преджелудках сохранилась до 4 месяцев и дальше к взрослому состоянию сайгаков (рис. 3). Очень интересным представляется тот факт, что на всем протяжении онтогенеза динамика гликогена в преджелудках была почти синхронной, при этом большие его значения были в книжке, средние – в сетке и наименьшие – в рубце, что коррелирует с интенсивностью процессов гистогенеза в этих камерах и является одним из адаптивных механизмов, обеспечивающих поддержание энергетического баланса в период морфогенеза.
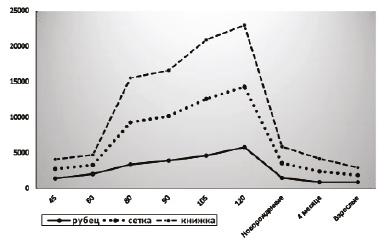
Рис. 3. Динамика гликогена в преджелудках сайгаков в течение онтогенеза
Гистохимическими и биохимическими методами выявлено, что начиная с раннеплодного периода и до взрослого состояния эпителий преджелудков сайгаков обладал также высокой ферментативной активностью. В 45 дней на поверхности эпителия преджелудков сайгаков выявились липаза, сукцинатдегидрогеназа, щелочная и кислая фосфатазы. На протяжении раннеплодного периода активность всех ферментов усиливалась. Локализовались они преимущественно в базальной мембране, клеточных оболочках, стенках кровеносных сосудов и нервных клетках.
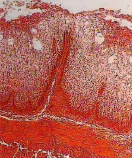
К концу позднеплодного периода в эпителии преджелудков увеличивалась активность липазы, сукцинатдегидрогеназы, щелочной и кислой фосфатаз (рис. 4). Это, очевидно, способствовало активации участия слизистой оболочки преджелудков в обменных процессах, через двусторонний транспорт метаболитов через эпителиальный пласт. Эти данные также коррелируют с особенностями гистогенеза камбиального слоя эпителия всех преджелудков, связанными с образованием особого базального комплекса за счет увеличения складчатости проксимальной поверхности базального слоя, наличием между пузырчатыми эпителиоцитами множества щелей и микроворсинок.

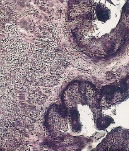

а) б) в)
Рис. 4. Локализация ферментов в стенках преджелудков поздних плодов сайгаков: а) рубец, щелочная фосфатаза, б) сетка, липаза, в) книжка, кислая фосфатаза. Ув. х200
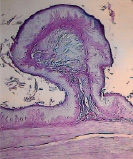
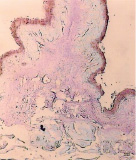
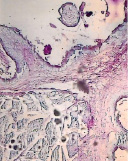
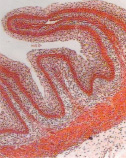
а) б) в)
Рис. 5. Гликокаликс на поверхности эпителиального пласта преджелудков взрослых сайгаков: а) рубец, б) сетка, в) книжка. Ув. х200
Гистохимическими методами в преджелудках плодов сайгаков в 45 дней были выявлены также нейтральные, а в 70 дней – кислые гликопротеины (рис. 5).
Известно, что гликопротеины участвуют в обменных процессах, ферментативных реакциях, росте, регенерации и выработке защитных свойств организма. На поверхности эпителия преджелудков гликопротеины, вместе с углеводами и ферментами, формируют гликокаликс – мощный защитный слой, увеличивающийся к рождению и до взрослого состояния сайгаков (рис. 5).
Заключение
Таким образом, в результате проведенных исследований выявленные морфофункциональные особенности преджелудков сайгаков в течение онтогенеза позволили выяснить развитие адаптивных механизмов к условиям обитания. Формирование адаптивных механизмов начинается уже в начале раннеплодного периода и продолжается с совершенствованием на протяжении всего онтогенеза. Морфологические особенности преджелудков заключались в основном в строении эпителиального пласта с большим количеством вакуолей, образованием особой складчатости проксимальной поверхности базальных эпителиоцитов, наличием межклеточных щелей и микроворсинок, что обеспечивало защитные свойства и двусторонний транспорт веществ, а также пристеночный и внутрипластовый метаболизм, которые усиливались к рождению, достигая завершенности у взрослых животных.
Функциональные особенности заключались в развитии обменных процессов, в преджелудках сайгаков в течение онтогенеза, осуществлявшихся в эмбриональный период через межуточный обмен с помощью двустороннего транспорта веществ через эпителий, направленной на поддержание постоянства внутренней среды организма. О транспорте веществ свидетельствовала и активность ферментов: липазы, сукцинатдегидрогеназы, щелочной и кислой фосфатаз, локализующихся в базальной мембране, по клеточным границам эпителиального пласта, в эндотелии кровеносных сосудов и нервных клетках. К рождению экскреторная функция слизистой оболочки преджелудков уменьшалась, вакуоли в эпителии постепенно исчезали, чему способствовало и включение амниотрофного питания в позднеплодный период путем заглатывания амниотической жидкости. У взрослых сайгаков обмен веществ в преджелудках усиливался, осуществляясь через межклеточные щели эпителиального пласта, обеспечивая сохранение энергетического баланса всего организма.
Выявленные механизмы морфологических и функциональных адаптаций преджелудков в процессе индивидуального развития сайгаков свидетельствуют о их значимости уже с начала их закладки и свидетельствуют о выраженности типа пищеварения жвачных, сформировавшегося и закрепившегося в течение длительной эволюции. Очевидно, обнаруженные механизмы экологически детерминированы, что говорит о необходимости сохранения условий обитания диких видов жвачных копытных, резкое изменение которых вследствие негативного антропогенного пресса может привести к непоправимым последствиям.
Полученные нами данные уточняют и расширяют представления о раннем морфогенезе желудка, формировании адаптивных механизмов диких жвачных копытных, что имеет не только теоретическое, но и практическое значение для разработки основ рационального кормления при разведении и содержании их в неволе с целью сохранения биоразнообразия.
Библиографическая ссылка
Хацаева Р.М. РАЗВИТИЕ МОРФОФУНКЦИОНАЛЬНЫХ МЕХАНИЗМОВ АДАПТАЦИИ ПРЕДЖЕЛУДКОВ САЙГАКОВ В ОНТОГЕНЕЗЕ К УСЛОВИЯМ ОБИТАНИЯ // Международный журнал прикладных и фундаментальных исследований. 2018. № 9. С. 115-120;URL: https://applied-research.ru/en/article/view?id=12397 (дата обращения: 03.07.2026).
DOI: https://doi.org/10.17513/mjpfi.12397