
Scientific journal
International Journal of Applied and fundamental research
ISSN 1996-3955
ИФ РИНЦ = 0,556
EXPLORATORY BEHAVIOR OF THE BANK VOLE (CLETHRIONOMYS GLAREOLUS) IN DIFFERENT PHASES OF POPULATION DYNAMICS

Ориентация представляет собой постоянный фоновый признак в жизни любого животного. Она возникает в ответ на новую или необычную форму стимуляции. Одним из основных свойств ориентировочной реакции является то, что ее возникновение связано с новизной раздражителя, и в то же время выраженность этой реакции зависит от видовой принадлежности [1, 2] и предыдущих условий жизни [3]. Кроме этого, на основании селекционных экспериментов было показано существование генетической компоненты в структуре исследовательского поведения грызунов [4, 5]. Все это свидетельствует о важной роли ориентировочно-исследовательского поведения в микроэволюционных процессах. Цель исследований состояла в изучении ориентировочно-исследовательского поведения рыжей полевки на разных фазах динамики численности.
Материалы и методы исследования
Рыжая полевка (Clethrionomys glareolus) – фоновый вид лесов Южного Урала. В течение трех полевых сезонов ежемесячно с апреля по сентябрь на контрольном участке в 3,5 га проводили живоотлов и мечение рыжих полевок. Численность грызунов за период исследований испытывала значительные колебания (рис. 1). Первый год мониторинга характеризовался очень низкой численностью. Весенняя численность рыжей полевки составляла 2,6 ос/га, осенняя – 9,7 ос/га (рис. 1; I – фаза депрессии). Весной следующего года численность составила 6,0 ос/га, осенью – 16,9 ос/га (рис. 1; II – фаза подъема численности). В течение третьего сезона наблюдений весенняя численность составляла 10,6 ос/га, осенью плотность рыжей полевки достигла максимальной величины, составив 65,1 ос/га (рис. 1; III – фаза пика численности). В этот сезон размножение грызунов прекратилось рано, уже в конце июля. Для изучения поведения отловленных зверьков приносили в полевой виварий. Перед экспериментом каждого зверька выдерживали в отдельной клетке не менее суток. Ориентировочно-исследовательское поведение грызунов изучали, используя модифицированный тест «открытое поле». Размер вольеры для наблюдений составлял 145х145х50 см. Пол был покрыт серым полимиктовым песком, который менялся перед каждым новым экспериментом. В целом условия были менее стрессирующими, чем в экспериментах с использованием классического «открытого поля», что, на наш взгляд, давало больше возможности наблюдать естественное поведение и процессы, происходящие в природных условиях. Эксперименты проводили во временном интервале от 18 час. 30 мин. до 24 час. Все наблюдаемое поведение грызунов регистрировалось в течение 10 минут. Важную роль в ориентации и исследовательской активности грызунов играет обоняние и зрение, поэтому к ориентировочным реакциям относили вертикальные стойки и оглядывание, принюхивание и обнюхивание пола. Зимовавших зверьков тестировали в конце апреля – начале мая, сеголеток – в августе, когда численность была максимальна. Из всех прибылых зверьков использовали только сеголеток последних генераций. Возраст полевок определяли по весу, а также благодаря мечению. После тестирования грызунов выпускали обратно в точке вылова. Всего в экспериментах было использовано 152 рыжих полевки (88 самцов и 64 самки). Ориентировочно-исследовательское поведение оценивали по следующим параметрам: общее время, затраченное на визуальные реакции, общее время, затраченное на обнюхивание пола, латентный период первого обнюхивания пола, латентный период первой визуальной ориентации. При изучении поведения индивидуальные различия часто маскируют анализируемые закономерности. Исключение или ослабление этого фактора обычно осуществляется путем увеличения числа подопытных животных с последующей статистической обработкой. Однако этот путь не всегда дает желаемый эффект. В нашем исследовании мы изначально были ограничены рамками одной микропопуляции и не имели возможности увеличения количества тестируемых зверьков. Преодолеть барьер индивидуальной изменчивости нам удалось за счет неоднократного тестирования каждого животного. Оценку различий в поведении проводили, используя критерий Стъюдента, характер взаимосвязей между переменными определяли с помощью корреляционного анализа.
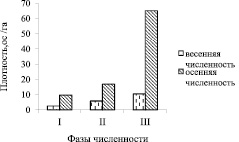
Рис. 1. Изменение плотности рыжей полевки на контрольном участке: I – фаза депрессии; II – фаза подъема численности; III – пик численности
Результаты исследования и их обсуждение
Исследовательское поведение, вызываемое неизвестной обстановкой и новыми предметами, представлено у грызунов поведенческими актами и позами, которые направлены на сбор информации о незнакомой ситуации, при этом выраженность ориентировочных реакций может иметь существенную изменчивость. Выявление размаха изменчивости поведенческих признаков может служить показателем состояния животного и способствовать идентификации факторов влияния в окружающей среде.
Коэффициенты вариации (Кв) общего времени ориентировочных реакций (ОВОР), определенные по суммарной выборке за все годы, составили в группе зимовавших – у самцов 100,4 %, самок – 90,8 %, сеголеток, соответственно – 53,8 % и 63,9 %.
У самцов сеголеток в фазу подъема величина ОВОР повышена, в фазу депрессии и фазу пика численности – снижена (рис. 2). Достоверные различия обнаружены между фазой подъема и фазой пика численности (p < 0,05). В группах сеголеток между годами не обнаружено достоверных различий. Кв по годам у сеголеток составляли: в фазу депрессии у самцов – 60,7 %, самок – 53,0 %, в фазу подъема соответственно – 47,0 % и 63,1 %, в фазу пика численности – 58,8 % и 79,4 %. Достоверных половых различий по величине ОВОР у зимовавших зверьков не обнаружено, хотя в фазу подъема тенденция к различиям есть.
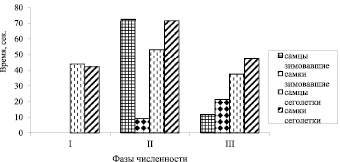
Рис. 2. Изменение общего времени ориентировочных реакций (ОВОР) рыжей полевки на разных фазах динамики численности. Обозначения такие же, как на рис. 1
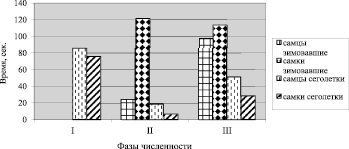
Рис. 3. Изменение латентного периода первой визуальной ориентации (ЛПВР) рыжей полевки на разных фазах динамики численности. Обозначения такие же, как на рис. 1
Сравнение зимовавших самцов и сеголеток показывает, что в целом продолжительность визуальных ориентаций у сеголеток больше, а изменчивость по годам меньше. У самок, так же как у самцов, величина ОВОР больше у сеголеток, чем у зимовавших. Различия достоверны в фазу подъема (p < 0,05).
Латентный период первой визуальной ориентации. Известно, что между стимулом и реакцией существует скрытый, или латентный период, обусловленный задержкой проведения импульсов через синапсы, и чем сильнее стимул, тем короче латентный период [6]. Сложные формы поведения, как и рефлексы, также имеют латентные периоды, и они не менее четкие, чем у рефлексов, но измерять их труднее, так как зачастую не удается точно фиксировать момент начала действия стимула на животное. Сложность определения латентных периодов заключается еще и в том, что на поведение могут влиять и другие факторы, изменяющие пороги появления реакции. Один из путей преодоления всех этих трудностей – использование большого числа животных, а затем сравнение средних значений латентных периодов реакций. Как и в случае рефлексов, наиболее сильные стимулы вызывают реакцию с наименьшим латентным периодом. Имеющиеся данные по поведению в «открытом поле» стрессированных и нестрессированных крыс свидетельствуют, что хроническое, вызванное в эксперименте, стрессирование животных увеличивает латентный период, т.е. на величину латентных периодов влияют общее состояние возбудимости нервной системы и чувствительность сенсорных систем к восприятию специфических сигналов [6].
Кв латентного периода первой визуальной ориентации (ЛПВР) по суммарной выборке составил у зимовавших самцов 120,6 %, зимовавших самок – 146,4 %, сеголеток соответственно: 135,5 % и 142,1 %.
У самок сеголеток и самцов сеголеток достоверная разность по величине ЛПВР обнаружена между фазой депрессии и фазой подъема (p < 0,05; рис. 3). Кв по годам для ЛПВР у сеголеток имели следующие значения: в фазу депрессии у самцов 132,8 %, самок – 72,3 %, в фазу подъема соответственно – 71,5 % и 64,2 %, фазу пика – 152,2 % и 138,5 %. У зимовавших самцов обнаружены достоверные отличия по величине ЛПВР между фазой подъема и фазой пика (p < 0,05). У зимовавших самок длительность латентного периода в течение двух лет оставалась практически одинаковой (рис. 3). Разность по величине ЛПВР между зимовавшими самцами и самками в фазу подъема была равна 96,9 с (разность не достоверна, но тенденция к различиям есть); в фазу подъема она сократилась до 16,3 с, в основном за счет длительности латентного периода у самцов.
Ольфакторное поведение. Ольфакторным поведением обычно называют элементы поведения, связанные с обонянием. Основной критерий ольфакторного поведения – это продолжительность реакции животного на источник запаха, регистрируя который можно охарактеризировать обоняние животного на уровне целостного организма с учетом всех его биологических и в первую очередь экологических особенностей. С помощью химических (обонятельных) сигналов животные осуществляют обмен информацией. Установлено, что все сложные формы поведения подвержены влиянию обонятельных сигналов, т.е. запахи влияют не только на отдельные органы, но и на весь организм в целом, который отвечает на воздействие определенной двигательной реакции. Обонятельное поведение тесно связано с маркировочным, к которому относят практически любое оставление животным запаха. Многие животные используют запаховые метки для установления границ занимаемой территории и распознания особей своего вида. На запаховой информации основывается пространственная структура и внутрипопуляционные отношения. Ольфакторные реакции также играют большую роль в межвидовых коммуникациях в качестве изолирующих механизмов [7].
Кв общего времени, затраченного на обнюхивание земли (ОВОЗ) по суммарной многолетней выборке составил: у зимовавших самцов – 86,6 %, зимовавших самок – 78,6 %, сеголеток – 47,0 % и 58,4 %.
Сравнение величин ОВОЗ показывает, что существенных половых и возрастных различий нет. В фазу депрессии у сеголеток разность в средних значениях ОВОЗ между самцами и самками составляла всего 2,9 с, причем длительность принюхиваний была больше у самцов, т.е. величины ОВОЗ у самок и самцов в фазу депрессии были практически одинаковыми. Однако в фазу подъема зимовавшие зверьки уже значительно различались: средняя длительность ольфакторных реакций у них была на 73,4 с больше, чем у самок (p < 0,05; рис. 4).
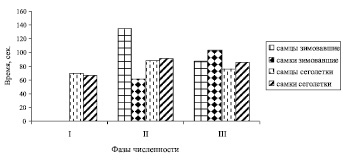
Рис. 4. Изменение общего времени, затраченного на обнюхивание земли (ОВОЗ) рыжей полевки на разных фазах динамики численности. Обозначения такие же, как на рис. 1
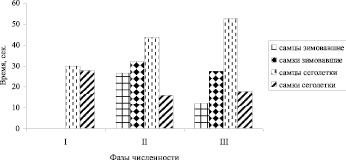
Рис. 5. Изменение латентного периода первого принюхивания (ЛППП) рыжей полевки на разных фазах динамики численности. Обозначения такие же, как на рис. 1
Латентный период первого принюхивания (ЛППП). Кв ЛППП по суммарным выборкам составляла у зимовавших самцов – 127,4 %, зимовавших самок – 127,5 %; в группе сеголеток, соответственно: 154,9 % и 115,6 %. Достоверные отличия по величине ЛППП обнаружены между зимовавшими самцами и зимовавшими самками (p < 0,05), а также между самцами сеголетками и зимовавшими (p < 0,01). Колебания величины ЛППП по годам более значительные у самцов. Сравнение по годам зимовавших самцов и сеголеток показало, что длительность латентного периода больше у сеголеток. Минимальная разница в средних, равная 16,7 с, отмечена в фазу подъема численности, максимальная (и достоверная, p < 0,05) – 40,7 с – в фазу пика численности (рис. 5).
Заключение
Результаты нашего исследования однозначно свидетельствуют, что между сеголетками и зимовавшими зверьками существуют различия по поведению, как в отдельные годы, так и по многолетним выборкам. Параметры поведения испытывают колебания по годам и зависят от уровня численности. В фазу подъема численности во всех половозрастных группах была увеличена длительность визуальных ориентаций и снижена их латентность (исключение составили зимовавшие самки, у которых длительность ориентаций была низкой, а их латентность высокая); у зимовавших самцов была увеличена длительность обнюхивания земли и латентность этой реакции, у зимовавших самок латентность этих реакций также была высокая. В фазу пика у самцов сеголеток коэффициент вариации латентного периода первой визуальной ориентации выше, чем в фазу депрессии. Такая же тенденция прослеживается у самок сеголеток. Поведение самцов и самок различается лишь количественно.
Библиографическая ссылка
Киселева Н.В. ОРИЕНТИРОВОЧНО-ИССЛЕДОВАТЕЛЬСКОЕ ПОВЕДЕНИЕ РЫЖЕЙ ПОЛЕВКИ (CLETHRIONOMYS GLAREOLUS) НА РАЗНЫХ ФАЗАХ ДИНАМИКИ ЧИСЛЕННОСТИ // Международный журнал прикладных и фундаментальных исследований. 2018. № 11-1. С. 94-98;URL: https://applied-research.ru/en/article/view?id=12456 (дата обращения: 04.07.2026).