
Scientific journal
International Journal of Applied and fundamental research
ISSN 1996-3955
ИФ РИНЦ = 0,556
RESISTANCE OF TREES AND SHRUBS IN FOREST-AGRARIAN LANDSCAPES OF DRY STEPPES

Вековой опыт создания лесных насаждений в засушливых районах европейской территории России, говорит что состояние значительной части защитных лесных насаждений в регионе нельзя признать удовлетворительным. Проведенные многочисленные исследования позволили предположить, что причинами их недостаточной долговечности являются различия между видовыми потребностями в обеспечении влагой и питательными веществами и фактическими условиями произрастания [1]. Нарушения, допущенные при составлении ассортимента, а также отклонение от научно обоснованной технологии выращивания защитных лесных насаждений в экстремальных условиях [2–4].
Экстремальные факторы по-разному влияют на жизнеспособность древесных видов. Поэтому их биологию следует учитывать при создании разнотипных защитных лесных насаждений.
Целью работы является оценка особенности роста и развития выявления механизмов эволюции и адаптации интродуцированных древесных пород в аридных условиях засушливого Нижнего Поволжья. Одновременно оценивается морозоустойчивость видов при предварительном воздействии экстремальных засушливых условий.
Материалы и методы исследования
В конкретных почвенно-климатических условиях оценка генофонда проводится в основном на трех группах объектов: аборигенного (естественного), искусственного (защитные лесные насаждения и культуры), насаждения и культуры, заложенные для исследовательских целей представителями различных опытных и научных учреждений. Морозоустойчивость выделенных деревьев определялась по обмерзанию побегов и сохранности после исключительно суровых зим. В холодильных установках дополнительно проводилось искусственное промораживание 1–2-летних потомств с ненарушенной корневой системой. Поврежденные растения учитывались путем окрашивания тканей солями тетразолия по истечении 1–2 суток. При этом температура, при которой погибало 50 % растений, считалась критической. Ткани древесных пород имеют разную физиологическую значимость. Это потребовало введения условных коэффициентов и общей их оценки в условных единицах. Значения коэффициентов: для камбия – 10, для флоэмы – 4, для ксилемы – 4, и для сердцевины – 2. В итоге давалась общая оценка в условных единицах.
Результаты исследования и их обсуждение
В условиях Нижнего Поволжья (г. Волгоград) нами выявлено при проведении исследований в регионе, что продолжительность роста побегов у дуба составляет от 19 до 24 суток, и он приходится на начало мая (табл. 1).
Таблица 1
Особенности роста побегов у дуба в онтогенезе в г. Волгограде
|
Показатели |
10–13 лет |
36–39 лет |
||||
|
1992 |
1994 |
1995 |
1992 |
1994 |
1995 |
|
|
Первый прирост |
||||||
|
Продолжительность роста, дней |
19 |
24 |
20 |
19 |
22 |
19 |
|
Прирост, см: |
||||||
|
верхушечный |
26,4 |
28,6 |
31,3 |
6,8 |
5,3 |
4,8 |
|
в т.ч. за сутки |
1,3 |
1,2 |
1,6 |
0,3 |
0,2 |
0,3 |
|
Боковой |
11,6 |
13,4 |
16,8 |
16,7 |
12,6 |
12,2 |
|
в т.ч. за сутки |
0,6 |
0,6 |
0,8 |
0,8 |
0,6 |
0,6 |
|
Второй прирост |
||||||
|
Продолжительность роста, дней |
14 |
20 |
13 |
– |
18 |
– |
|
Прирост, см: |
– |
– |
||||
|
верхушечный |
16,5 |
24,8 |
17,4 |
– |
6,4 |
– |
|
Боковой |
10,2 |
11,4 |
10,8 |
– |
16,8 |
– |
|
Количество растений с двумя приростами, % |
32 |
68 |
16 |
– |
7 |
– |
Скорость роста побегов зависит от целого ряда факторов, важнейшими среди которых являются погодные условия и возраст растений. В среднем в сутки побеги дуба растут от незначительного 0,2 до 1,9 см. При этом в условиях недостатка влаги суммарная величина годичного прироста обычно невелика. Растения теряют способность к образованию нескольких приростов. По многолетним наблюдениям число дубков со вторыми побегами в 10–13-летнем возрасте составило 68 %, в 36–39-летнем – 7 %.
Отмечено, что в сухой степи дуб развивается по скороспелому типу с максимальными приростами в 10–15 лет (рис. 1), в отличие от его развития в естественном ареале на Украине и Северном Кавказе, когда самые большие приросты наблюдаются в возрасте 30–40 лет [5].
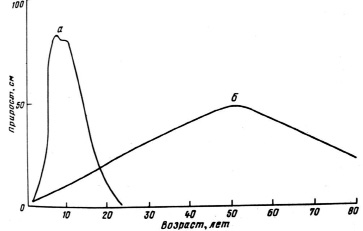
Рис. 1. Кривая прироста дуба по высоте: а) на каштановых почвах Ергеней; б) в естественных дубравах
В процессе эволюции при дифференциации жизненных процессов устойчивость явилась основополагающим фактором.
Толерантность и гомеостаз древесных видов в аридных условиях произрастания отображает их способность сохранять климаксные функции при аддитивном воздействии экстремальных факторов. Некоторые породы, обладающие способностью хорошо переносить морозы, могут легко повреждаться высокими летними температурами. Следовательно, генетическая структура биотипа является определяющим фактором его устойчивости организмов и индивидуальна [6].
Организм сначала приспосабливается к отдельным факторам, действующим на него, а уже затем, в процессе эволюции, к их комплексу. Селекционными мероприятиями можно изменить эффект воздействия того или иного фактора, повысить толерантность вида и биотипа и его долговечность.
Визуально физиологическое состояние дерева определяется по его кроне. В молодом возрасте кроны деревьев в основном островершинные и конусные в более старшем за счет активного роста боковых побегов. Усиленный рост верхушечного побега, образование островершинной конусновидной кроны в молодом возрасте сменяется снижением прироста верхушечного побега, возрастанием роста боковых ветвей они становятся ширококонусовидными, шаровидными и потом зонтиковидными.
Число меристемных клеток, способных к делению, увеличивается в период максимального роста, в более старшем возрасте эта способность снижается, что происходит одновременно с общим падением прироста. «Продуктивность камбия» – показатель, характеризующий способность клеток к миозу, – отношение прироста древесины по объему за год к площади поверхности камбиальной ткани [3].
Происходящие в тканях возрастные изменения вызывают падение его продуктивности.
В «период физиологической старости» дерева при частом повторении засух продуктивность камбия деградирует до состояния, когда растения не отзываются на улучшение адиотических условий [2, 3].
Отмечается быстрое прохождение онтогенеза в засушливых условиях, раннее плодоношение, старение и гибель, что может быть вызвано комплексом приспособительных реакций на резкое ухудшение гидрологических условий. Общая биологическая толерантность растения напрямую связана с его засухоустойчивостью [2–4].
Приход и расход определяет водный баланс древесных пород. Растения в процессе эволюции адаптации к почвенной засухе сокращают листовую поверхность, частично или полностью сбрасывают листву в период засухи [1, 2].
В отношении засухоустойчивости древесных видов можно предположить, что разные растения, даже одного вида не отличаются одинаковой потребностью к абиотическим факторам и под влиянием внешней среды могут в известных пределах адаптироваться [2, 6].
Одной из самых распространенных реакций растений на недостаток влаги в сухой степи является редукция листовой пластинки. Как и для всех ксерофитных растений, признаками, которые постепенно приобретаются в процессе эволюции, являются наличие водоносной и запасающей тканей, толстая кутикула, наличие в клетках слизи, масел, кристаллов, утолщение стенок эпидермиса, развитие столбчатой паренхимы, общая мелкоклеточность и плотность тканей.
В экстремальных условиях роста у листьев устойчивых древесных пород поддерживается на высоком уровне обмен веществ, увеличивается число сосудов в проводящих системах, и они значительно приближаются друг, к другу собираясь в своеобразный пучок, одновременно повышается вязкость протоплазмы, что позволяет сохранять тургор и избежать ожогов у листьев дуба, вяза приземистого и граболистного, ясеня ланцетного, робинии лжеакации даже в экстремальные засухи 1972, 1975 и 2010 гг. [2].
Низкая температура – абиотический фактор, значительно снижающий долговечность защитных лесных насаждений в аридной зоне. Вымерзание корневых систем в малоснежные зимы часто приводит к гибели древесных растений [2]. Правильный подбор и районирование древесных видов в данном случае является основным мероприятием против вымерзания [7]. Необходимо подбирать древесные породы с определенным «запасом морозоустойчивости», обеспечивающим их сохранность при абсолютных минимальных температурах данного региона. Для сохранения ассортимента древесных пород в условиях минимальных температур предложено выделить температуру, при которых погибает около 50 % растений, и назвать их критическими – сложное динамическое свойство, складывающееся из многих компонентов, так как в природных условиях на растения действует много факторов во время всего периода вегетации и среди них нелегко выделить основные. В процессе селекционной оценки растений на морозоустойчивость и зимостойкость трудно выделить основные факторы среди массы влияющих на этот процесс [2, 4]. Поэтому решили моделировать экстремальную обстановку путем прямого промораживания в климакамере. Установлено, что у ясеня ланцетного и саксаула черного низкие температуры до –16… –18 °С в период зимнего покоя не вызывают отпада, частичное повреждение отмечено у дуба и робинии при температуре –10…–12 °С; а при –13 –14 °С отпад более 50 % растений, полное вымерзание наблюдалось при температурах –18…–19 °С. При температурах –20…–24 °С и ниже происходит гибель растений ясеня, клена и саксаула (табл. 2).
Таблица 2
Аддитивное влияние факторов среды на морозоустойчивость древесных пород, г. Волгоград
|
Виды |
Сохранность ( %) после искусственного промораживания растений, предварительно выращенных в условиях |
||||||||||||
|
Оптимума |
Засухи |
Засоления |
|||||||||||
|
–8 ° |
–10 ° |
–12 ° |
–14 ° |
–16 ° |
–18 ° |
–20 ° |
–22 ° |
–8 ° |
–10 ° |
–12 ° |
–8 ° |
–10 ° |
|
|
Дуб черешчатый |
100 |
96 |
50 |
50 |
30 |
6 |
0 |
0 |
64 |
40 |
18 |
0 |
0 |
|
Ясень зеленый |
100 |
100 |
100 |
100 |
100 |
100 |
100 |
84 |
100 |
100 |
100 |
67 |
9 |
|
Клен ясенелистный |
100 |
100 |
100 |
100 |
100 |
96 |
89 |
80 |
100 |
90 |
80 |
0 |
0 |
|
Робиния лжеакация |
100 |
91 |
82 |
43 |
40 |
0 |
0 |
0 |
73 |
47 |
80 |
35 |
18 |
|
Саксаул черный |
100 |
100 |
100 |
100 |
100 |
87 |
– |
– |
100 |
100 |
100 |
87 |
58 |
Морозоустойчивость достигает максимума в январе и снижается к концу зимы у ясеня ланцетного, дуба черешчатого, клена ясенелистного, робинии лжеакации, саксаула черного в течение осенне-зимнего периода является величиной непостоянной и напрямую зависит от изменения температуры воздуха и почвы (табл. 3).
Таблица 3
Динамика морозоустойчивости древесных пород в осенне-зимний период
|
Виды |
Температуры ( °С), при которых гибнет 50 % растений по срокам промораживания |
||
|
декабрь |
январь |
февраль – март |
|
|
Ясень ланцетный |
–18…–20 |
–25 и ниже |
–13…–14 |
|
Дуб черешчатый |
–10…–11 |
–14…–16 |
–7…–8 |
|
Клен ясенелистный |
–18…–20 |
–24 и ниже |
–14…–15 |
|
Робиния лжеакация |
–13…–14 |
–13…–14 |
–7…–8 |
|
Саксаул черный |
–14…–14 |
–22…–24 |
–10…–11 |
Ясень ланцетный показал более высокую устойчивость к морозу, чем дуб черешчатый и робиния.
Периодические засухи, предшествующие суровым зимам, также снижают морозоустойчивость древесных видов. После глубокой засухи при благоприятном водном режиме почвы и достаточно теплой погоде происходит возобновление роста побегов у робинии лжеакации, что значительно ослабляет растение и снижает его морозоустойчивость. У ясеня ланцетного, клена ясенелистного, дуба черешчатого и саксаула черного подобные процессы не отмечались. Вследствие чего, робиния лжеакация после наступления низких температур имела отпад при снижении температры до –8 °С. У других видов морозоустойчивость не показала такой прямой зависимости от засухи.
Изучение зависимости влияния экологических факторов позволяет комплексно оценить адаптивные возможности разных видов и рекомендовать их для культивирования в определенных почвенно-климатических условиях.
Выводы
1. Древесные растения в аридной зоне развиваются по ускоренному типу, они раньше вступают в стадию плодоношения, более интенсивно плодоносят, хотя качество семян обычно невысокое. У них более высокая активность камбия и интенсивный рост по диаметру. Раньше на 20–30 % наступает старение и гибель, чем в зоне экологического оптимума.
2. Моделированием экстремальной обстановки путем прямого промораживания в климакамере установлены критические низкие температуры для основных пород защитных лесных насаждений.
3. Засухи перед экстремальными зимами значительно снижают морозоустойчивость древесных видов. Растения ослаблены, не успевают подготовиться в экстремальный летний период и сильнее, чем обычно, страдают от воздействия низких зимних температур.
Библиографическая ссылка
Иозус А.П., Завьялов А.А., Крючков С.Н. УСТОЙЧИВОСТЬ ДЕРЕВЬЕВ И КУСТАРНИКОВ В ЛЕСОАГРАРНЫХ ЛАНДШАФТАХ СУХОЙ СТЕПИ // Международный журнал прикладных и фундаментальных исследований. 2018. № 11-1. С. 124-128;URL: https://applied-research.ru/en/article/view?id=12462 (дата обращения: 21.07.2026).