
Scientific journal
International Journal of Applied and fundamental research
ISSN 1996-3955
ИФ РИНЦ = 0,556
THE ROLE OF ANNELID WORMS (ANNELIDA) IN THE NAROCH LAKES SYSTEM (BELARUS)

Изучение изменений, вызванных различными факторами, и происходящих в процессе функционирования озерных экосистем, является одной из задач лимнологии [1]. Решение вопроса об эволюционных процессах в экосистемах приводит, в том числе, к необходимости изучения биоразнообразия водоемов и экологических условий, которые лежат в основе его формирования [2]. Кольчатые черви Annelida (Clitellata: Oligochaeta, Hirudinea) относятся к одной из широко распространенных в разнообразных условиях, географических широтах и типах водоемов групп макрозообентоса. Однако, несмотря на их значение в динамике водных экосистем [3, 4], исследования биологии и роли в оценке качества вод [5–8], знания об их распределении и экологии, по сравнению с другими группами бентоса, достаточно ограничены. Интерес к изучению Annelida в озерах Нарочанской системы (Беларусь) обусловлен тем фактом, что эти озера прошли несколько этапов эволюции [9, 10]. Это показано как на примере физико-химических параметров среды, так и на примере развития различных элементов биоты [10, 11]. Наблюдение за развитием одной из преобладающих групп зообентоса в исследуемых озерах позволит не только расширить сведения об экологии данной группы беспозвоночных, но и выявить вероятные закономерности развития червей в озерах различного трофического статуса.
Цель исследования: анализ данных о развитии одной из групп макрозообентоса (Annelida) в озерах с разным трофическим статусом.
Материалы и методы исследования
В анализ вошли данные, полученные сотрудниками НИЛ гидроэкологии и Нарочанской биологической станции БГУ (Беларусь) за время регулярных мониторинговых наблюдений на озерах Нарочанской системы в период с 1997 по 2016 г., а также использованы архивные материалы, частично представленные в работе [12]. Отбор проб макрозообентоса в оз. Нарочь, Мястро и Баторино проводили на постоянных станциях по принципу полуразрезов от берега к наибольшей глубине (от 5 до 8 точек), в течение вегетационного сезона с мая по октябрь. Подробное описание отбора гидробиологических проб зообентоса приведено в работе [12]. Численность и биомассу донных животных пересчитывали на площадь дна (1 м2) в зависимости от их распределения по исследованным глубинам в соответствии с морфометрическими данными озер. В настоящей работе обсуждается средняя взвешенная численность и биомасса бентоса. Наличие и оценку степени связи между признаками определяли с помощью коэффициента корреляции Пирсона. Перед расчетом коэффициентов корреляции показатели подвергли логарифмированию по основанию 10.
Результаты исследования и их обсуждение
В настоящей работе мы рассматриваем следующие этапы: 1) антропогенного эвтрофирования (1978–1983 гг.); 2) олиготрофикации (с 1984 по 1990 г. для оз. Нарочь и с 1984 по 1989 г. для озер Мястро и Баторино); 3) период бентификации (с 1991 г. для оз. Нарочь и с 1990 г. для озер Мястро и Баторино по настоящее время). Все они характеризуются определенными параметрами среды и уровнем развития биоты [11]. На современном этапе, который описывается как период бентификации, каждое из трех озер каскада имеет свой уровень трофности: Нарочь (олиго-мезотрофный), Мястро (мезотрофный) и Баторино (эвтрофный водоем). Отмечается, что продуктивность озер в этот период возросла за счет роли донных и прикрепленных сообществ [10, 11].
В составе донных сообществ Нарочанских озер группа аннелид играет заметную роль. С момента начала регулярных мониторинговых наблюдений в 1997 г. олигохеты встречены в половине, а пиявки в 30 % гидробиологических проб. В общей численности бентоса доля олигохет составляла от 19,1 до 35,9 %, в биомассе – от 10,6 до 15,8 %. Эти же значения для пиявок были несколько ниже: по численности они составляли 0,04–5,1 %, а по биомассе – 0,06–16,3 %.
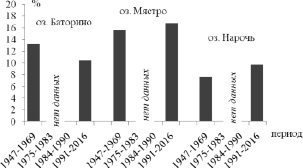
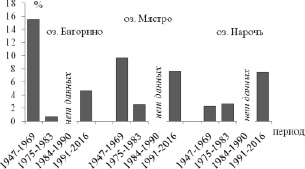
Анализ количественных данных развития олигохет по временным периодам (рис. 1) показал, что период бентификации (с 1991 г. по сегодняшний день) характеризуется снижением доли олигохет в общей численности и биомассе бентоса, по сравнению с другими периодами, в эвтрофном оз. Баторино и повышением этих показателей в олиготрофном оз. Нарочь.
А)
Б)
Рис. 1. Доля ( %) численности (А) и биомассы (Б) олигохет от общей биомассы и численности зообентоса в озерах Нарочанской системы
С 1991 по 2016 г. наибольшие средние значения численности и биомассы олигохет отмечаются в мезотрофном озере Мястро (253,8 ± 50,1 экз/м2 и 0,62 ± 0,12 г/м2) и олиготрофном озере Нарочь (235,4 ± ± 12,9 экз/м2 и 0,93 ± 0,06 г/м2). Наименьшие показатели количественного развития олигохет (73,5 ± 18,1 экз/м2 и 0,09 ± 0,02 г/м2) указывались для эвтрофного озера Баторино. Так же в оз. Нарочь и Мястро отмечались наибольшая численность (74,6 ± 9,6 и 21,4 ± 7,0 экз/м2) и биомасса (1,16 ± 0,19 и 0,22 ± 0,07 г/м2) пиявок в сравнении с оз. Баторино (8,0 ± 6,1 экз/м2 и 0,03 ± 0,01 г/м2). Сходное соотношение параметров на современном этапе развития озер отмечалось в целом и для общего бентоса: наибольшие значения численности и биомассы указывались для озер Нарочь и Мястро [12].
Известно, что все изменения характеризующие этапы развития озерных экосистем сопровождаются структурными перестройками сообществ, как различных звеньев трофической цепи [13, 14]. Для анализа зависимости показателей развития олигохет и факторов среды были выбраны параметры, изменяющиеся в разные периоды и указывающие на изменение трофического статуса озер [11]: прозрачность воды (SD), концентрация азота (Ntot) и фосфора (Ptot), углерода (Ctot) содержание взвешенного вещества (sest), концентрация хлорофилла (Chl), биохимическое потребление кислорода за 5 сут. (БПК5), электропроводность (cond). Для отображения трофического статуса озер использовался индекс трофического состояния Карлсона (TSI).
Результаты расчета коэффициента корреляции (таблица) показали, что количественные показатели развития олигохет имеют сильную прямую взаимосвязь (p = 0,001) только с прозрачностью воды, а с остальными – обратную.
Коэффициенты корреляции Пирсона между количественными показателями развития олигохет (N oligoch – численность, экз/м2; B oligoch – биомасса, г/м2), индексом Карлсона и гидроэкологическими параметрами Нарочанских озер
|
SD |
TSI |
sest |
БПК5 |
Ctot |
Ntot |
Ptot |
Chl |
cond |
|
|
N oligoch |
0,62* |
–0,55* |
–0,62* |
–0,51* |
–0,58* |
–0,53* |
–0,54* |
–0,63* |
|
|
B oligoch |
0,77* |
–0,72* |
–0,78* |
–0,66* |
–0,74* |
–0,60* |
–0,52* |
–0,72* |
–0,68* |
Примечание. * p ≤ 0,001.
Все указанные параметры коррелируют между собой и индексом TSI [15], поскольку входят в формулу его расчета. Таким образом, на фоне снижения значений индекса TSI, концентрации биогенных элементов, изменения гидрологических показателей озер увеличиваются показатели количественного развития олигохет (рис. 2).
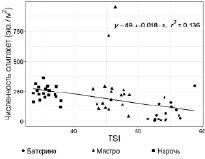
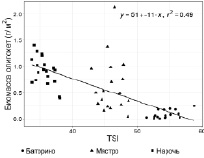
Рис. 2. Связь TSI с численностью, экз/м2 и биомассой, г/м2 олигохет в Нарочанских озерах. Коэффициенты корреляции приведены в таблице
Заключение
Регулярный учет зообентоса, начиная с 1997 г. по современный период, который приходится на этап бентификации в развитии Нарочанских озер, дает представление о динамике количественных показателей развития групп донных беспозвоночных в озерах разного трофического статуса. Численность и биомасса аннелид (Oligochaeta и Hirudinea), достигает более 40 % и 30 % общего бентоса соответственно. Положительная динамика развития кольчатых червей в Нарочанских озерах может быть связана со снижением трофического статуса озер и, соответственно, с параметрами, определяющими этот статус. Дополнительную информацию о водной системе, ее изменениях и структуре макрозообентоса может дать идентификация таксонов до видового уровня. Это подтверждает необходимость проведения дальнейших таксономических исследований этой группы донных беспозвоночных.
Работа выполнена при поддержке РФФИ в рамках научного проекта № 18-54-00009 Бел_а и гранта Белорусского республиканского фонда фундаментальных исследований Б18P – 056.
Библиографическая ссылка
Батурина М.А., Макаревич О.А., Кайгородова И.А., Жукова Т.В., Адамович Б.В. РОЛЬ АННЕЛИД (ANNELIDA) В ОЗЕРАХ НАРОЧАНСКОЙ СИСТЕМЫ (БЕЛАРУСЬ) // Международный журнал прикладных и фундаментальных исследований. 2018. № 12-1. С. 56-59;URL: https://applied-research.ru/en/article/view?id=12521 (дата обращения: 01.07.2026).
DOI: https://doi.org/10.17513/mjpfi.12521