
Scientific journal
International Journal of Applied and fundamental research
ISSN 1996-3955
ИФ РИНЦ = 0,556
INTERACTION OF COMPONENTS OF THE ANTIOXIDANT SYSTEM IN CEREALS DURING DROUGHT

Абиотические факторы и неблагоприятные климатические условия приводят к стрессу, сопровождающемуся рядом метаболических изменений у растений. Одним из главных факторов стресса является засуха. Проблема глобального потепления может привести к тому, что климат станет еще более засушливым. Поэтому изучение механизмов адаптации растений к засухе и на сегодняшний день остается актуальным.
Цель наших исследований заключалась в изучении активности компонентов антиоксидантной защитной системы у образцов различных разновидностей и сортов злаковых культур при воздействии засухи.
Материалы и методы исследования
В качестве материала исследования использовались образцы пяти разновидностей мягкой пшеницы (Triticum aestivum L.): var. delfi k-79, var. erythrospermum k-10, var. hostianum k-50, var. albidum k-25, var. velutinum k-30 и трех сортов ячменя: Pallidum 596, Polongi, Flor. Растения выращивались в условиях фитотрона при температуре 20–21 °C, влажности 60–70 %, 16-часовом фотопериоде, освещенности 10000 люкс. Пятидневные проростки в течение 24 ч подвергались воздействию умеренной и сильной засухи, моделированной с помощью осмотических растворов сахарозы в 14 и 20 атм. Определялись активность гваякол-пероксидазы и содержание пролина в листьях [1, 2].
Определение активности пероксидазы спектрофотометрическим путем основано на измерении оптической плотности продуктов, образующихся в ходе реакции окисления гваякола. После измельчения листьев (200 мг) в фарфоровой чашке с небольшим количеством (5-10 мл) фосфатного буфера (рН 5,4) смесь центрифугировали в течение 10 минут со скоростью 4000-5000 об/мин. Оптическая плотность реакционной смеси, состоящей из 0,5 мл H2O2, 0,5 мл субстрата (гваякола), 1,5 мл фосфатного буфера, 0,5 мл супернатанта (ферментативного растительного материала), измеряли на спектрофотометре (УФ-3100 ПК) в течение 1 мин, при длине волны 470 нм. Оптическую плотность пролина определяли при длине волны 520 нм.
Результаты исследования и их обсуждение
Исследование образцов мягкой пшеницы и ячменя показало, что во время засухи активность гваякол-пероксидазы и содержание пролина в листьях всех образцов значительно менялись относительно контроля.
Мягкая пшеница. В контрольных вариантах активность фермента менялась в диапазоне 0,111–0,243, имея максимальный показатель у проростков var. delfi k-79, а минимум у var. erythrospermum k-10 (рис. 1).
Во время умеренной засухи (14 aтм) активность фермента варьировала в диапазоне 0,232–0,367, а при воздействии сильной засухи (20 aтм) – между 0,087 и 0,234. Минимум активности фермента был отмечен у образцов var. albidum k-25 как при среднем стрессе, так и при высокой концентрации сахарозы (соответственно, 0,232 и 0,087), а максимум – у проростков var. hostianum k-50 (0,367) при умеренной, и у растений var. erythrospermum k-10 (0,234) при сильной засухе.
Образцы, участвующие в эксперименте, продемонстрировали различные изменения в процентном отношении к контролю. Так, во время средней засухи в 14 aтм наблюдалось увеличение активности гваякол-пероксидазы во всех вариантах опыта. Наибольшее увеличение было отмечено у проростков var. erythrospermum k-10 (223,0 %) и var. hostianum k-50 (207,3 %).
При увеличении силы воздействия стресса (20 aтм) наблюдали частичное или резкое ослабление активности фермента: самый высокий показатель все еще был отмечен у растений var. erythrospermum k-10 (210,8 %). Активность гваякол-пероксидазы у образцов var. delfi k-79, var. albidum k-25, var. velutinum k-30 и var. hostianum k-50 снизилась и по отношению к контролю составила 36,6; 44,6; 57,6 и 65,5 % соответственно. В течение суток активность фермента в проростках var. delfi k-79 снизилась на 0,154 единицы, что составило 36,6 % от контроля, что является суммой максимального расхода конституционного пула пероксидазы в течение дня. Ослабление активности фермента, вызванное сильной засухой, можно объяснить большим количеством повреждений в растениях, расходом фермента, большим по сравнению с его синтезом, его деградацией, или увеличением активности низкомолекулярных компонентов защитной системы, выполняющих в клетке антиоксидантную функцию [3].
Содержание свободного пролина в контроле менялось в пределах 0,083–0,12 µМ/мг. Минимальный показатель наблюдался у растений var. erythrospermum k-10, максимальный у var. albidum k-25 (рис. 2).
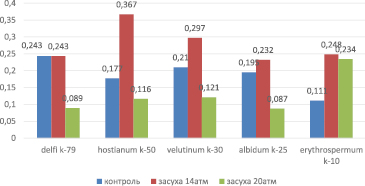
Рис. 1. Активность гваякол-пероксидазы у образцов мягкой пшеницы в условиях умеренной и сильной засухи (14 и 20 aтм)
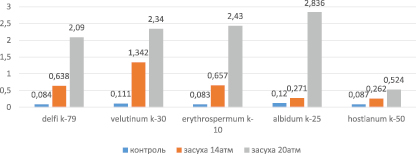
Рис. 2. Cодержание пролина в образцах мягкой пшеницы (µM/мг) в условиях умеренной (14 aтм) и сильной (20 aтм) засухи
Во всех вариантах опыта наблюдалось увеличение содержания этой аминокислоты. При умеренном воздействии стресса (14 атм) содержание пролина менялось в интервале 0,262–1,342 µM/мг. У образцов var. hostianum k-50 отмечены минимальные, а у var. velutinum k-30 –максимальные значения содержания аминокислоты. В то время как у var. albidum k-25 содержание пролина увеличилось более чем в 2,25 раза, а у var. hostianum k-50 – в 3 раза по отношению к контролю, показатели var. velutinum k-30 были выше в 12,09 раза. Влияние сильной засухи (20 атм) также несколько увеличило количество аминокислоты – в 6–23,6 раза по сравнению с контролем. Самый низкий уровень наблюдался у образцов var. hostianum k-50, а самый высокий у экспериментального варианта var. albidum k-25. В целом содержание пролина изменялось в диапазоне 0,524–2,836 µM/мг. Предельные значения относились к опытным вариантам var. hostianum k-50 и var. albidum k-25 соответственно.
У образцов var. hostianum k-50 влияние умеренной засухи привело к увеличению содержания свободного пролина на 0,175 µM/мг в течение суток, в то время как при воздействии сильной – прирост составлял 0,437 µM/мг относительно контроля. В проростках var. albidum k-25 эти цифры составили 0,151 µM/мг и 2,716 µM/мг соответственно. То есть антиоксидантная система защиты усилила синтез низкомолекулярного компонента в зависимости от силы воздействия стресса. Однако резкий рост содержания пролина у растений var. albidum k-25 не исключает того, что, наряду с синтезом, количество пролина может увеличиваться за счет деградации белков [3].
Повышение дозы стрессового фактора, а в нашем случае засухи, приводит к большему увеличению содержания свободного пролина в растительной ткани. Поскольку пролин обладает осмопротекторной и антиоксидантной функциями, следовательно, значительное увеличение его содержания направлено на защиту растения от резкого воздействия стресса.
Ячмень. Аналогичная тенденция наблюдалась и у ячменя. У контрольных растений содержание свободного пролина менялось в диапазоне 0,096–0,16 µM/мг (рис. 3). Образцы Pallidum 596 имели минимальные показатели, а опытный вариант Polongi – максимальные. Влияние засухи средней степени (14 aтм) не вызвало большого увеличения содержания аминокислоты, оно достигало 0,127–0,170 µM/мг. Это свидетельствует о том, что ячмень устойчив к умеренной засухе. Аналогичные данные были получены и в работах других исследователей [4].
Влияние стрессора высокой интенсивности (20 атм) привело к значительному увеличению содержания свободного пролина, значения которого варьировали в диапазоне 0,227–0,310 µM/мг. Максимальное содержание аминокислоты наблюдалось у сорта Polongi, а минимальное – у сорта Pallidum 596. Однако по отношению к контролю содержание пролина у сорта Polongi увеличилось в 1,93 раза; у сорта Pallidum 596 – в 2,36 раза; у сорта Flor – в 2,46 раза.
Активность ферментативного компонента антиоксидантной системы защиты, т.е гваякол-пероксидазы, находилась в обратно пропорциональной зависимости с содержанием низкомолекулярного компонента – пролина. Так, активность гваякол-пероксидазы при стрессе средней интенсивности (14 атм) значительно возросла и изменилась в интервале 0,571–0,675 (рис. 4). У контрольных растений этот показатель варьировал между 0,123 и 0,144. Максимальное увеличение активности фермента наблюдалось у опытного варианта сорта Flor – в 8,13 раза больше контрольного, минимальное – у растений сорта Polongi, что в 3,96 раза превосходит контрольный вариант. Активность гваякол-пероксидазы в опытном варианте Pallidum 596, повышаясь, составила 0,606, что в 4,26 раза выше, чем в контроле.
При воздействии засухи высокой интенсивности (20 aтм) активность пероксидазы изменялась в диапазоне 0,138–0,338. У образцов сорта ячменя Pallidum 596 активность фермента снизилась до 0,138, что составило 97,0 % от контрольного показателя. Активность фермента в вариантах опыта Polongi и Flor увеличилась и, соответственно, составила 0,338 (в 2,35 раза больше, чем в контроле) и 0,310 (в 2,25 раза больше, чем в контроле).
Следует отметить, что независимо от вида зерновой культуры, при воздействии умеренной и сильной засухи наблюдалась обратно-пропорциональная зависимость между содержанием пролина и активностью гваякол-пероксидазы. Так, если во время сильного стресса, по сравнению с умеренным, содержание пролина у образцов var. delfi k-79 увеличилось в 3,2 раза, то активность фермента ослабла в 2,7 раза. Эти показатели составляют 1,7 и 3,1 у var. velutinum k-30; 3,6 и 2,5 у var. erythrospermum k-10; 2,0 и 1,1 у var. hostianum k-50 соответственно. У образцов var. albidum k-25 содержание пролина при сильном стрессе по сравнению с умеренным значительно возросло и увеличилось в 10,4 раза, а активность пероксидазы ослабла в 2,6 раза.
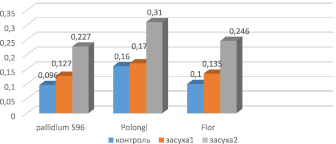
Рис. 3. Содержание пролина при стрессе у растений ячменя: 1 – контроль; 2 – умеренная засуха (14 aтм); 3 – сильная засуха (20 aтм)
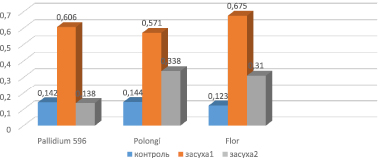
Рис. 4. Активность пероксидазы у образцов ячменя в условиях стресса: 1 – контроль; 2 – умеренная засуха (14 aтм); 3 – сильная засуха (20 aтм)
Незначительное увеличение содержания пролина сопровождалось значительным повышением активности гваякол-пероксидазы в ходе воздействия стресса умеренной интенсивности у всех трех сортов ячменя. А при воздействии сильной засухи, напротив, значительное увеличение содержания свободного пролина сопровождалось умеренным увеличением активности фермента. Так, по сравнению с засухой средней интенсивности, при сильной засухе содержание свободного пролина в проростках ячменя сорта Polongi увеличилось в 1,9 раза, а активность пероксидазы снизилась в 1,7 раза. Если у образцов сорта Flor содержание пролина увеличивалось в 1,8 раза, то активность фермента уменьшалась в 2,1 раза. У проростков сорта Pallidum 596 содержание пролина увеличилось в 1,8 раза, однако активность пероксидазы снизилась даже ниже значения контроля, что в 4,39 раза ниже, чем при воздействии умеренного стресса. Полученные результаты свидетельствуют о взаимодействии компонентов системы антиоксидантной защиты.
Согласно представленным многочисленным экспериментальным данным, изменение активности антиоксидантных ферментов зависит от дозы, продолжительности действия стресс – фактора, уровня активности ферментов и генотипа растения, используемого в исследовании [5]. Но эффективность работы антиоксидантных ферментов не всегда достаточна для детоксикации свободных радикалов, образующихся в больших количествах при усилении воздействия стресса [6]. Результаты свидетельствуют о том, что пролин несет антиоксидантную функцию и участвует в гашении свободных радикалов. Кроме того, пролин играет важную роль в поддержании клеточного метаболизма, обеспечивая выживание растений в экстремальных условиях [7]. С этой точки зрения выявленные нашими экспериментами различия в активности ферментативных и низкомолекулярных компонентов антиоксидантной системы у пшеницы и ячменя вносят определенный вклад в изучение механизма защиты растений при стрессе. Таким образом, если значительное повышение активности фермента гваякол-пероксидазы при воздействии стресса умеренной интенсивности (14 aтм) на проростки исследуемых образцов пшеницы и ячменя сопровождалось незначительным увеличением содержания аминокислоты пролина, то в условиях высокой дозы (20 aтм) ферментативная защитная система значительно снижала свою активность, а низкомолекулярный компонент антиоксидантной системы, в данном случае пролин, активировался, что подтверждается многократным увеличением его содержания в проростках. На основании этих данных можно утверждать, что во время засухи происходит взаимодействие ферментативных и низкомолекулярных компонентов антиоксидантной системы защиты в тканях растительного организма, т.е. можно говорить о наличии реципрокных отношений. В ряде литературных источников также есть информация о возможном наличии реципрокной связи между пролином и антиоксидантными ферментами. Подобная закономерность была обнаружена у ряда дикорастущих растений [8] и пшеницы [9].
Библиографическая ссылка
Ибрагимова З.Ш., Меджидова Г.С., Гасанова Г.И., Абдуллаева Л.С., Алиев Р.Т. ВЗАИМОДЕЙСТВИЕ КОМПОНЕНТОВ АНТИОКСИДАНТНОЙ СИСТЕМЫ У ЗЛАКОВЫХ КУЛЬТУР ПРИ ЗАСУХЕ // Международный журнал прикладных и фундаментальных исследований. 2020. № 9. С. 7-11;URL: https://applied-research.ru/en/article/view?id=13119 (дата обращения: 25.05.2026).