
Scientific journal
International Journal of Applied and fundamental research
ISSN 1996-3955
ИФ РИНЦ = 0,556
GEROPROTECTIVE POTENTIAL OF CHRONOBIOTIC KL001 IN VIVO AND IN SILICO

Группа экспериментальных препаратов, называемых хронобиотиками, представляет собой структурно разнородную совокупность малых молекул, показавших в экспериментах высокую эффективность в отношении циркадного ритма экспрессии генов на модельных объектах или в культуре. В список мишеней хронобиотиков главным образом входят именно белки-компоненты молекулярного механизма клеточных часов, поддерживающих осцилляции циркадного ритма. Среди хронобиотиков следует выделить два типа, прямые и опосредованные, и несколько наиболее крупных классов в зависимости от мишени внутри первого типа: лиганды казеин киназ, лиганды криптохромов, лиганды ROR-рецепторов, соединения, связывающие REV-ERB и ингибиторы GSK-3 [1, 2].
Ингибиторы казеин киназ представлены лонгдейзином, DK359, NCC007, PF4800567, эпибластинами A and C (лиганды CK I) [1] и DMAT, TBB, CX-4945, GO289 (лиганды CK II) [1]. GSK-3-специфичные ингибиторы представлены CHIR99021, BRD1652. Интеракторы рецепторов RORs представлены в основном соединениями-агонистами: нобилетином, нейрорускогенином, T0901317, SR1001, SR2211, SR1555, SR3335 и SR1078. Подгруппа активаторов криптохрома включает KL001, Compound 50, KL044, GO200, KL101, TH301 и др., в то время как в подгруппу ингибиторов входят KS15-дериваты и неэтоксипропаноевые производные [2, 3].
Непрямые хронобиотики гораздо более многочисленны и разнородны, отличает их от прямых то, что для них не установлена конкретная мишень (компонент клеточных часов), которая модифицирует циркадный ритм при связывании с молекулой-лигандом. Среди непрямых хронобиотиков можно выделить модуляторы фототрансдукторного каскада (опсинамиды и ингибиторы меланопсина), неспецифичные ингибиторы GSK-3 (литий и производные бензодиазипина), индукторы экспрессии Bmal1 (L-methyl selenocysteine), активаторы Sirt, такие как ресвератрол (спорная подгруппа, поскольку у млекопитающих Sirt вовлечен в осциллятор, как второстепенный элемент механизма; существуют также хронобиотики, для которых мишени вообще не описаны, например «Compound 10/CEM3». Стоит отметить, что геропротекторный потенциал был описан лишь у лития и производных бензодиазепина [4, 5].
Активатор криптохрома был избран в качестве объекта исследования в силу его способности модифицировать экстраокулярную фоторецепцию. В качестве гипотезы мы выбрали утверждение, что стабилизация криптохрома дрозофилы dCRY продлевает жизнь плодовой мухи при сверхэкспрессии гена cry. Чтобы проверить гипотезу, решено было использовать KL001, активатор криптохрома [5]. Дрозофилиный криптохром dCRY был избран в качестве мишени из-за существования экспериментальных доказательств наличия геропротекторного эффекта генетических вмешательств в экспрессию гена cry [6].
Цель исследования: проверить способность KL001 модифицировать эффект продления жизни, вызванный пан-нейрональной конститутивной экспрессией гена cry у самцов дрозофилы; провести компьютерное моделирование взаимодействия белка-мишени dCRY c лигандом KL001.
Материалы и методы исследования
Линия y, w;UAS-cry24/CyO (трансген располагается на 2 хромосоме, присутствуют две копии) – обе конструкции являются несущими дополнительные копии гена cry (криптохром) в сопровождении промоторной последовательности UAS (upstream activating sequence). В эксперименте использовались линии дрозофил с выровненным в течение шести поколений генетическим фоном: линия UAS-cry24 и линия tim-GAL4-64 (генотип: w; tim-GAL4-64/+). Для выравнивания генетического фона использовали линию w1118. Самцов драйверной линии tim-GAL4 скрещивали с виргинными самками UAS-cry24 для получения особей, сверхэкспрессирующих ген cry в нервной системе. В качестве генетического контроля использовали особей родительских линий tim-GAL4 и UAS-cry24.
Климатические установки Binder KBF720-ICH (Binder, Германия) использовались для содержания экспериментальных животных. Температура поддерживалась постоянно на уровне 25 °С, относительная влажность воздуха составляла 60 %. В зависимости от варианта эксперимента использовалось два режима освещения: 12 ч свет и 12 ч темнота (нормальный), 24 ч темнота.
Особи из контрольной и опытных групп перемещались на свежую питательную среду одновременно, дважды за неделю до конца жизни. Навеска сухих компонентов питательной среды в расчете на 1 л воды: 30 г – сахарный песок, 30 г – манная крупа, 8 г – сухие дрожжи, 7 г – агар-агар по 8 мл – 50 % пропионовая кислота (водный раствор) и 10 % раствор нипагина в 96 %-ном этаноле.
Статистическая обработка результатов экспериментов по оценке выживаемости проводилась в приложении OASIS 2 [7].
Для изучения эффектов активатора криптохрома наносили 30 мкл раствора KL001 (Sigma-Aldrich, USA) в дистиллированной воде и 0,1 % DMSO в концентрациях 5 мкмоль/мл на поверхность питательной среды дрозофил. На среду контрольной группы животных наносили 30 мкл 0,1 % DMSO, в отрицательном контроле использовалась дистиллированная вода.
Анализ взаимодействий молекул рибофлавина, ФАД, KL001 и KS15 c криптохромом и ионом Mg2+ (pH = 7,4) проводили в графическом интерфейсе AmDock [8] на базе программ AutoDock Vina. Файл с координатами атомов криптохрома дрозофилы и иона Mg2+ был загружен из базы данных PDB под кодовым номером 4gu5.
Результаты исследования и их обсуждение
Данные, полученные при проведении эксперимента по подсчету продолжительности жизни самцов дрозофил, получавших KL001 в дозе 5 мкМоль/мл и одновременно экспрессирующих конструкт UAS-cry24/CyO / tim-GAL4-64 (пан-нейрональная конститутивная экспрессия криптохрома), показали, что препарат KL001 в сочетании с растворителем 0,1 % DMSOводн нивелирует геропротекторный эффект сверхэкспрессии криптохрома. В значительной степени в отрицательном контроле выражен эффект самой сверхэкспрессии, который воспроизводится относительно более ранних исследований, однако в опытной группе, которая получала все воздействия (препарат, растворитель, воду, сверхэкспрессию), статистически значимых различий с контролем, не сверхэкспрессировавшим криптохром, не обнаружено. Однако следует отметить, что KL001 в некоторой степени нивелирует отрицательное воздействие растворителя. Особи гибридного генотипа (UAS-cry24/CyO / tim-GAL4-64), получавшие препарат, по сравнению с особями контрольной группы, имевшими тот же генотип и получавшими DMSO, обнаруживают геропротекторный эффект (увеличение медианной продолжительности жизни на 34 %, p < 0,0001). Обнаруженный эффект стоит признать несущественным, поскольку при сравнении подопытной группы с отрицательным контролем (вода) по тому же генотипу UAS-cry24/tim-GAL4-64, наблюдается снижение медианной продолжительности жизни на 17 %. Таким образом, KL001 в присутствии растворителя, 0,1 % раствора DMSOводн, способен полностью отключать механизм продления жизни, опосредованный сверхэкспрессией криптохрома у дрозофил (табл. 1). Следует признать, что геропротекторный эффект KL001, показанный ранее на диком типе линии Canton-S, является следствием криптохром-независимых механизмов, так как KL001, вероятно, выступает ингибитором дрозофилиного криптохрома и инактивирует его [9]. Последнее утверждение мы попытались доказать методом молекулярного докинга in silico (табл. 2).
Таблица 1
Параметры продолжительности жизни самцов дрозофилы, сверхэкспрессировавших криптохром и получавших KL001 c пищей
|
№ варианта |
Генотип и воздействие, самцы |
N |
Параметры средней |
Процентили смертности |
||||
|
Средняя ПЖ |
Ст. ош. cредней |
25 % |
50 % |
75 % |
90 % |
|||
|
1. |
tim-GAL4-64 / tim-GAL4-64 + KL001 5 мкМоль/мл |
181 |
39,60 |
0,60 |
34 |
41 |
48 |
– |
|
2. |
UAS-cry24/CyO / tim-GAL4-64 + KL001 5 мкМоль/мл |
155 |
55,75 |
0,70 |
55 |
55*+ |
62 |
64 |
|
3. |
UAS-cry24/CyO / UAS-cry24/CyO + KL001 5 мкМоль/мл |
166 |
56,66 |
0,89 |
48 |
58 |
68 |
69 |
|
4. |
tim-GAL4-64 / tim-GAL4-64 + 0.1 % DMSOводн |
151 |
22,61 |
0,61 |
15 |
22 |
29 |
32 |
|
5. |
UAS-cry24/CyO / tim-GAL4-64 + 0.1 % DMSOводн |
163 |
39,64 |
0,78 |
34 |
41+# |
48 |
52 |
|
6. |
UAS-cry24/CyO / UAS-cry24/CyO + 0.1 % DMSOводн |
153 |
32,06 |
0,88 |
24 |
31 |
41 |
45 |
|
7. |
tim-GAL4-64 / tim-GAL4-64 + H2O |
190 |
40,66 |
0,93 |
39 |
45 |
48 |
51 |
|
8. |
UAS-cry24/CyO / tim-GAL4-64 + H2O |
171 |
66,57 |
1,30 |
55 |
66$ |
79 |
92 |
|
9. |
UAS-cry24/CyO / UAS-cry24/CyO + H2O |
214 |
58,07 |
1,02 |
50 |
60 |
67 |
78 |
Примечания: * – p < 0,0001 (сравнение вариантов 2 и 5; 2 и 8; согласно критерию Гехана – Бреслоу – Уилкоксона с поправкой Бонферрони); + – p < 0,001 (сравнение вариантов 2 и 1; 5 и 4; 5 и 6; согласно критерию Гехана – Бреслоу – Уилкоксона с поправкой Бонферрони); $ – p < 0,001 (сравнение вариантов 8 и 7; 8 и 9; согласно критерию Гехана – Бреслоу – Уилкоксона с поправкой Бонферрони); # – p < 0,0001 согласно критерию Гехана – Бреслоу – Уилкоксона с поправкой Бонферрони, сравниваются варианты 5 и 8.
Следует заключить, что наивысшей аффинностью при учете минимальной эффективной концентрации по отношению к дрозофилиному криптохрому обладает молекула KL001 (рис. 1), в силу максимального сходства с нативным (референсным) лигандом ФАД (рис. 2), второе место занимает соединение KS15 (рис. 3), ингибирующее криптохром дрозофилы в микромолярных концентрациях.
Низкая константа ингибирования и высокая аффинность KL001 (табл. 2) обнаруживают высокий ингибиторный потенциал соединения, что указывает на криптохром-независимые механизмы в качестве основной причины продления жизни самцов в более ранней работе [9].
Таблица 2
Сравнение результатов молекулярного докинга ФАД и хронобиотиков KL001 и KS15 c молекулой белка dCRY
|
ФАД (Референсный нативный лиганд-кофактор) |
KL001 |
KS15 |
||||||||||
|
№ |
Аффинность |
Ki |
Единицы измерения |
Эффективность лиганда |
Аффинность |
Ki |
Единицы измерения |
Эффективность лиганда |
Аффинность |
Ki |
Единицы измерения |
Эффективность лиганда |
|
1 |
-16,4 |
0,0 |
нмоль |
-0,31 |
-8,2 |
975,81 |
нмоль |
-0,29 |
-7,3 |
4,46 |
мкмоль |
-0,28 |
|
2 |
-16,4 |
0,0 |
нмоль |
-0,31 |
-8,1 |
1,16 |
мкмоль |
-0,29 |
-7,1 |
6,25 |
мкмоль |
-0,27 |
|
3 |
-16,2 |
0,0 |
нмоль |
-0,3 |
-7,9 |
1,62 |
мкмоль |
-0,28 |
-7,1 |
6,25 |
мкмоль |
-0,27 |
|
4 |
-16,2 |
0,0 |
нмоль |
-0,31 |
-7,9 |
1,62 |
мкмоль |
-0,28 |
-7,1 |
6,25 |
мкмоль |
-0,27 |
|
5 |
-16,0 |
0,0 |
нмоль |
-0,3 |
-7,8 |
1,92 |
мкмоль |
-0,28 |
-6,8 |
10,37 |
мкмоль |
-0,26 |
Примечание. Ki – константа ингибирования.
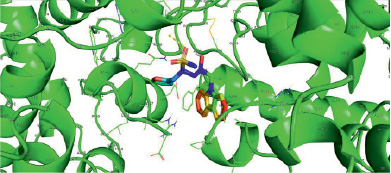
Рис. 1. KL001 взаимодействует с dCRY в кратере, связывающем ФАД
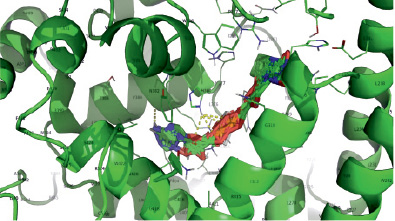
Рис. 2. Модель взаимодействия молекул ФАД с dCRY в энергетически наиболее выгодных позициях
Описано множество вариантов докинга ФАД, имеющих практически равную свободную энергию взаимодействия (от -16.0 до -16.4 ккал на рис. 3 представлены первые пять позиций из табл. 2).
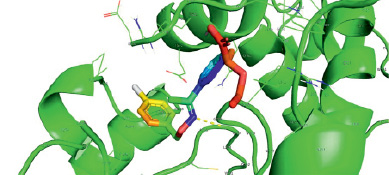
Рис. 3. KS15 взаимодействует с dCRY в кратере, связывающем ФАД
Заключение
Эффект продления жизни в условиях сверхэкспрессии гена, кодирующего мишень KL001, белок dCRY не сохраняется относительно отрицательного контроля (тот же генотип, особи получали воду с пищей), хотя в опыте относительно группы с тем же генотипом, подвергавшейся воздействию растворителя (0,1 % ДМСО) увеличение медианной продолжительности жизни наблюдалось существенное (+34 %, p < 0,0001).
1. Результаты моделирования взаимодействий белка dCRY c различными лигандами указывают на KL001 как на препарат, имеющий максимальное сродство с криптохромом дрозофилы (в сравнении с нативным кофактором ФАД), аффинность -8,2 ккал/моль, эффективная концентрация ингибирования 975,81 нмоль, эффективность лиганда – 0,29 (следует считать приемлемой).
2. Высокий ингибирующий потенциал KL001 свидетельствует о существовании криптохром-независимых механизмов продления жизни, индуцированных KL001 в более ранних экспериментах, так в настоящем исследовании эффект продления жизни самцов сверхэкспрессией криптохрома нивелируется употреблением экспериментальными животными в пищу KL001 в концентрации 5 мкМоль/мл (дозе, имеющей геропротекторные свойства).
Исследование выполнено при финансовой поддержке РФФИ в рамках научного проекта № 19-34-90058 «Аспиранты»: «Исследование геропротекторных свойств стабилизатора и ингибитора криптохрома, KL001 и KS15, на модели Drosophila melanogaster». Исследования выполнены в рамках государственного задания по теме «Разработка геропротекторных и радиопротекторных препаратов», № АААА-А19-119021590022-2.
Библиографическая ссылка
Соловьёв И.А., Шапошников М.В., Москалев А.А. ГЕРОПРОТЕКТОРНЫЙ ПОТЕНЦИАЛ ХРОНОБИОТИКА KL001 В IN VIVO ЭКСПЕРИМЕНТАХ И ЕГО ВАЛИДАЦИЯ IN SILICO // Международный журнал прикладных и фундаментальных исследований. 2021. № 9. С. 7-11;URL: https://applied-research.ru/en/article/view?id=13264 (дата обращения: 21.07.2026).