
Scientific journal
International Journal of Applied and fundamental research
ISSN 1996-3955
ИФ РИНЦ = 0,556
Study of the behavior of stochastic entropy in a 2D and 3D cellular automaton system

Введение
Клеточные автоматы (КА) более 50 лет применяются при моделировании сложных систем, где из локальных правил взаимодействия возникают пространственно-временные структуры [1, с. 21–52]. Теоретический анализ КА строится на теории информации: энтропия Шеннона количественно оценивает неопределенность дискретных состояний и их динамику [1, с. 237–240]. Методы изучения самоорганизации сформировались в классификациях правил КА [2, с. 22–25]. Характерный пример – тьюрингова универсальность «Игры в жизнь» Джона Конвея, показавшая вычислительную полноту простых КА [3, с. 89].
Круг задач, охватываемых современными исследованиями КА, достаточно широк. В частности, простейшие модели КА применяются при анализе динамических систем разной физической природы [4]. Примером могут служить методы решения классических уравнений математической физики на гексагональных сетках. Сначала их разработали для двух измерений [5], а затем распространили на трехмерные решетки. Результат – возможность учитывать пространственную анизотропию и граничные условия нестандартного типа [6]. Согласно обзору отечественных публикаций [7], интерес к стохастическим и асинхронным моделям КА растет, однако вопрос о влиянии размерности на информационную динамику проработан недостаточно.
Основное преимущество КА – способность порождать сложную макродинамику из локальных правил. Экспериментально показано, что на основе правил «Игры Жизни» можно строить логические элементы и ячейки памяти в дискретной среде [8]. Для количественной оценки эволюции таких систем широко используют энтропийный анализ. Классификация элементарных КА по энтропийным признакам позволяет выявлять переходы между упорядоченными, сложными и хаотическими режимами даже при асинхронном обновлении [9]. Стохастические эффекты и механизмы памяти в КА на данный момент находятся в центре внимания. Нейросетевые расширения КА, по имеющимся данным, эффективны в задачах обучения с подкреплением, но их энтропийные характеристики изучены слабо [10]. Предварительные данные авторов при сравнении 2D и 3D конфигураций в идентичных начальных условиях выявили выраженную дихотомию в энтропийной динамике; это требует более глубокого статистического обоснования [11]. В свою очередь, память в локальных правилах открывает возможности для конструирования сложных динамических режимов и управления переходами в распределенных системах [12]. Математическое и программное обеспечение стохастических КА с памятью уже разработано, с его помощью моделируют нелинейные эффекты коллективной синхронизации [13].
Дополнительное направление связано с представлением клеточных автоматов в матричной форме, когда состояние всей решетки обновляется как единая матрица или тензор, а правила задаются композициями линейных операций и поэлементных нелинейностей. Это позволяет использовать оптимизированные библиотеки тензорных вычислений и на порядки ускорять симуляции. В частности, Понтес-Фильо и соавт. [14] предложили общий нейро-вдохновленный фреймворк, в котором стохастические КА, случайные булевы сети и эхо-сети эволюционируют к критическому состоянию, а динамика эффективно реализуется матричными операциями. Такой подход открывает возможности для массового ансамблевого анализа энтропийных и других статистических характеристик крупномасштабных систем, что согласуется с целями настоящей работы.
В свою очередь, в трехмерных системах проблема обостряется. Трехмерные КА усложняют задачу экспоненциальным ростом числа конфигураций, затрудняя расчет энтропии [15]. Систематическое исследование трехмерных аналогов игры «Жизнь» было проведено Бейсом [16], выявившим множество правил и динамических режимов на кубических решетках. Эпидемиологические модели КА показывают зависимость динамики от размерности и топологии [15]. С точки зрения теории информации скорость роста энтропии в многомерных клеточных автоматах строго проанализирована Бургуэтом [17], который ввел понятие перенормированной энтропии и получил оценки для d-мерного случая. Кроме того, связь между кластерной структурой и перколяционными свойствами в вероятностных клеточных автоматах исследована Хартарски [18], что важно для понимания перехода от разреженных конфигураций к связным кластерам. Отсюда возникает необходимость систематически сравнить динамику стохастической энтропии и структурную организацию в 2D и 3D клеточных автоматах.
В работе проведено систематическое сравнение энтропии, фазовых траекторий и кластеров в 2D и 3D стохастических КА при ансамблевом усреднении.
Цель исследования – сравнение динамики стохастических клеточных автоматов 2D и 3D размерностей при идентичных начальных условиях и ансамблевом усреднении, а также выявить влияние размерности решетки на характер эволюции систем.
Материалы и методы исследования
Математические модели
В основе количественного анализа – энтропия Шеннона [1, с. 238]. Для стохастических клеточных автоматов она, как известно, выступает мерой неопределенности. Оценку проводят на двух уровнях: глобальном и локальном.
Сначала – глобальная энтропия. Пусть 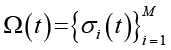 – конфигурация системы на временном шаге t, где M – общее число клеток в решетке, σi(t) ∈ {0,1} – бинарное состояние i-й клетки (1 – «живая», 0 – «мертвая»). Множество всех возможных глобальных конфигураций обозначим
– конфигурация системы на временном шаге t, где M – общее число клеток в решетке, σi(t) ∈ {0,1} – бинарное состояние i-й клетки (1 – «живая», 0 – «мертвая»). Множество всех возможных глобальных конфигураций обозначим
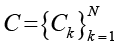 ,
,
причем полное число микросостояний N = 2M. Тогда глобальная стохастическая энтропия задается классической формулой
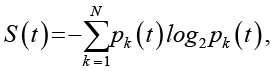 (1)
(1)
где pk(t) – вероятность реализации k-й конфигурации в момент t.
Прямое вычисление S(t) по формуле (1) для крупных решеток, например
503 = 125000 клеток,
невозможно, так как N растет экспоненциально. Выход: локальная оценка энтропии. Берем окрестность клетки (x, y, z) на шаге t. В двумерных задачах это окрестность Мура радиуса 1. Локальный паттерн N(x, y, z; t) – вектор состояний из Kc элементов, где Kc – число соседей (8 в 2D, 26 в 3D). Число уникальных конфигураций такой окрестности – 2Kc. При Kc = 26 получаем более 67 млн комбинаций.
Вместо полного перебора всех возможных паттернов применяется оценка частотного распределения локальных конфигураций по ансамблю симуляций. На каждом временном шаге t для каждой клетки (во всех реализациях) фиксируется локальный паттерн – состояния самой клетки и ее соседей в окрестности Мура (8 в 2D, 26 в 3D). Пусть {cj} – множество всех уникальных паттернов, встретившихся хотя бы один раз в ансамбле на шаге t, а Nobs(t) – их количество. Относительная частота fj(t) определяется как доля клеток во всем ансамбле, окрестность которых совпадает с паттерном cj:
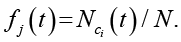 (2)
(2)
где Nci(t) – число клеток с паттерном ci в ансамбле на шаге t, N – общее число клеток во всех реализациях.
Тогда локальная энтропия для произвольной клетки, основанная на распределении паттернов по ансамблю, задается формулой
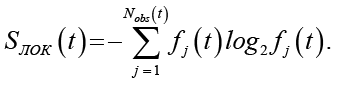 (3)
(3)
Данная величина не зависит от конкретной клетки и характеризует неопределенность локального микроскопического состояния, усредненную по системе.
Глобальную энтропию системы получают усреднением локальных значений по всем клеткам ансамбля; поскольку CЛОК одинакова для всех клеток, глобальная энтропия совпадает с (3):
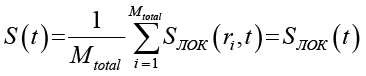 , (4)
, (4)
где ri – координаты i-й клетки. Mtotal – суммарное число клеток во всех реализациях ансамбля.
Описание параметров симуляций
Сравнительную серию экспериментов проводили для 2D и 3D клеточных автоматов. Решетки: квадратная 50×50 и кубическая 30×30×30. Выбрали их так, чтобы при разной размерности объем вычислений оставался сопоставимым, а кластерные структуры – репрезентативными. Граничные условия везде периодические – это исключает краевые эффекты.
Состояние каждой клетки бинарное: σi(t) ∈ {0,1}, единица соответствует «живой», ноль – «мертвой». Вероятность перехода в состояние 1 на следующем шаге задается логистической функцией от числа живых соседей ni(t) в окрестности Мура радиуса 1:
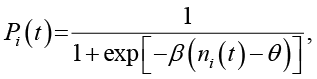 (4)
(4)
Параметр θ – порог активации, β – крутизна функции. Для 2D взяли θ = 4 (половина от 8 соседей), для 3D – θ = 12 (половина от 26). β = 2,0 в обоих случаях; это значение обеспечивает плавный, но достаточно резкий переход вблизи порога, что типично для моделей с умеренной стохастичностью.
Таблица 1
Параметры численных экспериментов для 2D и 3D КА
|
Параметр |
КА 2D |
КА 3D |
|
Размер сетки |
50×50 |
30×30×30 |
|
Тип решетки |
Квадратная |
Кубическая |
|
Граничные условия |
Периодические |
Периодические |
|
Окрестность (Мура) |
8 соседей |
26 соседей |
|
Начальная плотность |
0,25 |
0,25 |
|
Функция активации |
|
|
|
Режим обновления |
Частично асинхронный (α = 0,15) |
Полностью асинхронный + + коррелированный шум (σ = 0,05) |
|
Число независимых запусков |
50 |
50 |
|
Длительность симуляции |
1000 шагов |
1000 шагов |
|
Основные метрики |
Число живых клеток, ⟨S(t)⟩ |
Число живых клеток, ⟨S(t)⟩ |
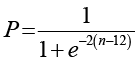
Примечание: составлена авторами на основе полученных данных в ходе исследования.
Начальная плотность живых клеток ρ0 = 0,25 одинакова для всех экспериментов. Актуальные параметры симуляций представлены в табл. 1.
Начальные конфигурации – случайные: для каждой клетки вероятность быть живой равнялась 0,25, реализация – через генератор NumPy. Чтобы сгладить влияние конкретного распределения, каждую модель прогоняли 50 раз; параметры θ, β и ρ0 при этом не менялись. Длительность прогона – 1000 шагов. Предварительные тесты показали: к этому моменту система либо выходит на квазистационарный режим (3D), либо достигает устойчивого состояния (2D).
Размеры решеток подбирали из соображений баланса, а именно: 50×50 для 2D и 30×30×30 для 3D – достаточно, чтобы наблюдать кластерные структуры, но не слишком велики, чтобы ансамбль из 50 запусков можно было выполнить за приемлемое время. Локальную энтропию считали по окрестности Мура: 8 соседей в 2D, 26 в 3D. Частотное распределение паттернов {fj(t)} оценивали по ансамблю; глобальную энтропию S(t) получали усреднением локальных значений по всем клеткам согласно (3). Дополнительно фиксировали число живых клеток, плотность, проводили кластерный анализ (связные компоненты по 8/26 соседям) и строили фазовые траектории. Реализация – на Python 3.11 в среде Google Colab. Для ускорения использовали компиляцию через Numba (декораторы @njit). Визуализация выполнена с помощью Matplotlib и Seaborn, статистическая обработка – SciPy и pandas.
Результаты исследования и их обсуждение
Результаты численного моделирования 2D и 3D стохастических клеточных автоматов показали: расхождения между размерностями не сводятся к количественным отличиям. Геометрия решетки и структура локальных взаимодействий порождают качественно разные типы динамики. Сравнение проводили по трем основным характеристикам: изменение численности популяции, эволюция энтропии Шеннона, форма фазовых траекторий.
Рисунок 1 отражает усредненные по 50 реализациям временные ряды числа живых клеток. В двумерном случае (Conway B3/S23, сетка 50×50) наблюдается выраженная релаксация: стартовая популяция 722 клетки снижается на 84,6 % и стабилизируется на уровне 112±15 клеток (4,4 % от площади). Высокий коэффициент вариации (CV составил 49,7 %) говорит о значительной стохастичности; формируются разреженные кластерные структуры. В трехмерной модели (Life B56/S4567, объем 30³) картина принципиально иная: среднее число живых клеток не только не падает, но даже немного возрастает относительно начального уровня (с 6763 до 6939) и стабилизируется вблизи значения 6939±47 клеток, что составляет примерно 25,7 % объема решетки. Коэффициент вариации числа живых клеток в стационарном режиме не превышает 0,68%, что на порядок ниже, чем в двумерном случае, и свидетельствует о высокой статистической устойчивости макроскопического состояния.
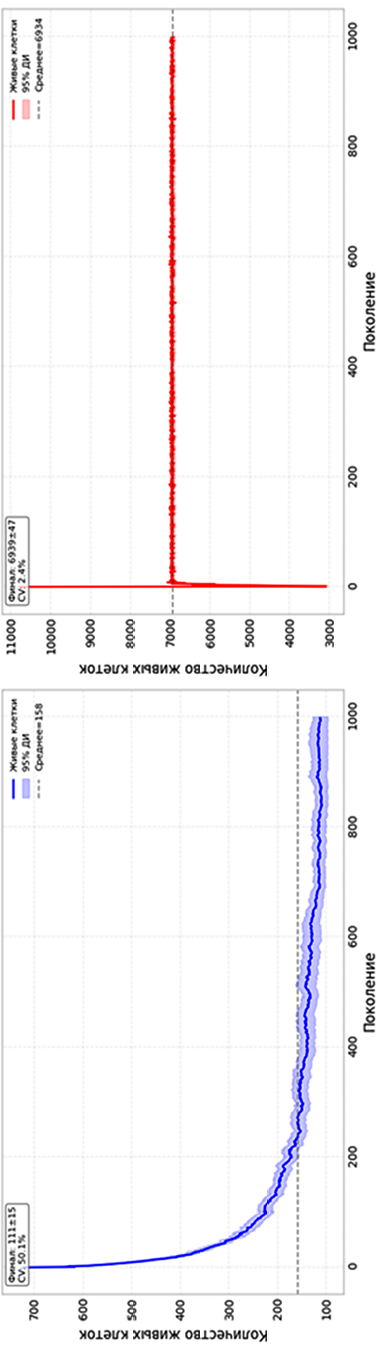
Рис. 1. Динамика числа живых клеток в КА 2D (а) и 3D (б) Примечание: составлен авторами по результатам данного исследования
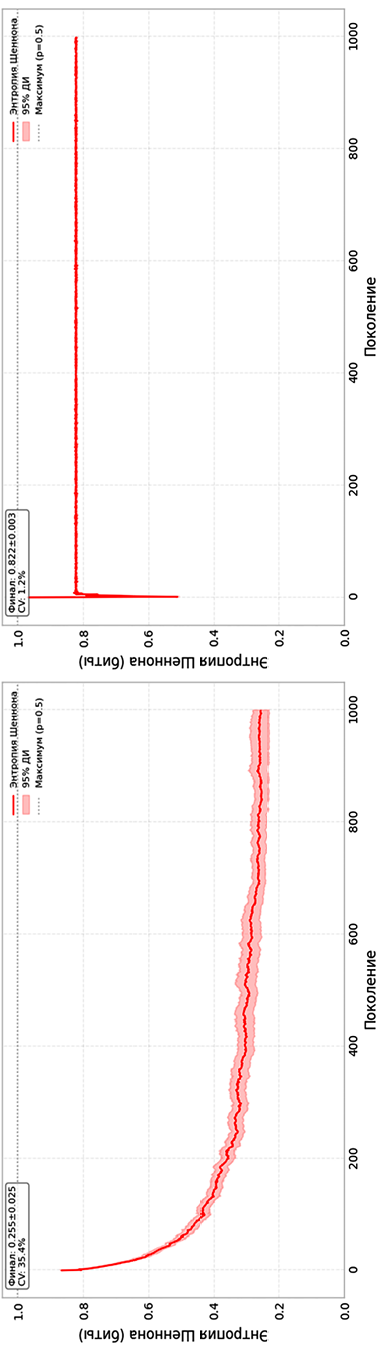
Рис. 2. Динамика стохастической энтропии в 2D (а) и 3D (б) КА Примечание: составлен авторами по результатам данного исследования
Поддерживается она значительной плотностью активных элементов и непрерывными локальными перестройками в пределах окрестности Мура.
Эволюция информационной энтропии Шеннона (рис. 2) демонстрирует контрастные режимы самоорганизации. Для двумерной системы характерен глубокий спад энтропии: от начального значения 0,866 бит к асимптотическому уровню 0,255±0,025 бит (снижение на 0,611 бит). Потеря ~71 % информационной емкости соответствует переходу в режим структурной редукции с формированием разреженных детерминированных паттернов. Коэффициент вариации энтропии в двумерной модели, оцененный как CV временного ряда средней энтропии ⟨S⟩ на последних 200 шагах, составил 35,4 %; CV самой финальной энтропии по ансамблю равен 35,3 % (табл. 2), оба показателя свидетельствуют о высокой остаточной изменчивости. Это довольно высокий показатель, и он говорит о том, что локальная динамика даже при общей тенденции к упорядочению остается стохастической. В трехмерной системе картина иная. Стационарный уровень энтропии здесь зафиксирован на отметке 0,822±0,003 бит, что составляет 82,2 % от максимально возможного значения (Smax). Снижение относительно начального – всего 0,149 бит. Оба коэффициента вариации энтропии в трехмерной системе минимальны: 1,2 % для временного ряда и 1,3 % для ансамблевого распределения (табл. 2). Разница между финальными значениями энтропии в 3D и 2D составляет 0,567 бит. Такое расхождение подтверждает, что режимы эволюции принципиально разные.
Столь высокая энтропия для 3D КА указывает на то, что микроскопические состояния системы остаются сильно неопределенными, а локальные паттерны не вырождаются в небольшой набор повторяющихся конфигураций. В рамках качественного анализа динамики клеточных автоматов такую ситуацию допустимо характеризовать как близкую к хаотическому состоянию, понимая под этим не строгую математическую хаотичность, а сохранение макроскопической неупорядоченности, сопоставимой со случайным распределением активных клеток. Об этом же свидетельствуют незатухающие флуктуации производной энтропии (рис. 3) и отсутствие сходимости фазовых траекторий к точечному аттрактору (рис. 4). Напротив, в двумерной модели энтропия падает до 0,255 бит – значения, характерного для сильно разреженных и частично детерминированных конфигураций. В этом случае доминирует процесс структурной редукции: информационная насыщенность снижается, а динамика приближается к режиму с выраженным преобладанием упорядоченных паттернов.
Производная энтропии по времени (dS/dt) показана на рис. 3. Ее значения позволяют судить о скорости релаксации информационных процессов. Для начального участка характерны отрицательные значения dS/dt. В трехмерной модели этот показатель равен −25,59±0,82×10⁻³ бит/поколение, в двумерной – −12,91±1,16×10⁻³ бит/поколение. В двумерной системе флуктуации dS/dt затухают. На последних шагах среднее значение составляет −0,048±0,581×10⁻³ бит/поколение, амплитуда колебаний σ = 1,71×10⁻³ бит/поколение. Это соответствует режиму, близкому к стационарному. В трехмерной системе картина иная. Колебания dS/dt не затухают: размах значений остается в интервале [−28,2; +35,5]×10⁻³ бит/поколение, амплитуда σ = 2,18×10⁻³ бит/поколение. Среднее значение за последние шаги равно −0,113±0,509×10⁻³ бит/поколение, то есть практически нуль, но знак колеблется.
Наблюдаемую динамику трехмерного клеточного автомата уместно охарактеризовать как метастабильную, понимая под этим следующее: на протяжении всего времени симуляции (1000 шагов) средняя энтропия ⟨S⟩ остается вблизи 0,822 бит без значимого тренда, коэффициент вариации энтропии на финальном участке не превышает 1,17 %, а амплитуда флуктуаций производной dS/dt составляет 2,18 × 10⁻³ бит/шаг и не затухает (табл. 2). При этом ни в одной из реализаций скользящее среднее энтропии не покидало полосу ±2σ относительно среднего уровня, то есть характерное время жизни метастабильного состояния превышает 1000 шагов. Такое сочетание макроскопической стабильности и микроскопических перестроек типично для метастабильных режимов в системах с локальными взаимодействиями.
Поскольку энтропия, будучи интегральной характеристикой, не позволяет судить о том, как именно организовано пространственное распределение активных элементов, дополнительно был проведен кластерный анализ с выделением связных компонент клеточного автомата. Соответствующие количественные оценки приведены в табл. 2. Для двумерной системы обращает на себя внимание кратное сокращение плотности кластеров – с 34,7 до 5,3, то есть более чем в 6 раз. Средний размер кластера при этом сокращается с 8,24 до 7,65 клетки, а корреляционная длина ξ падает с 1,72 до 1,18 ячейки.
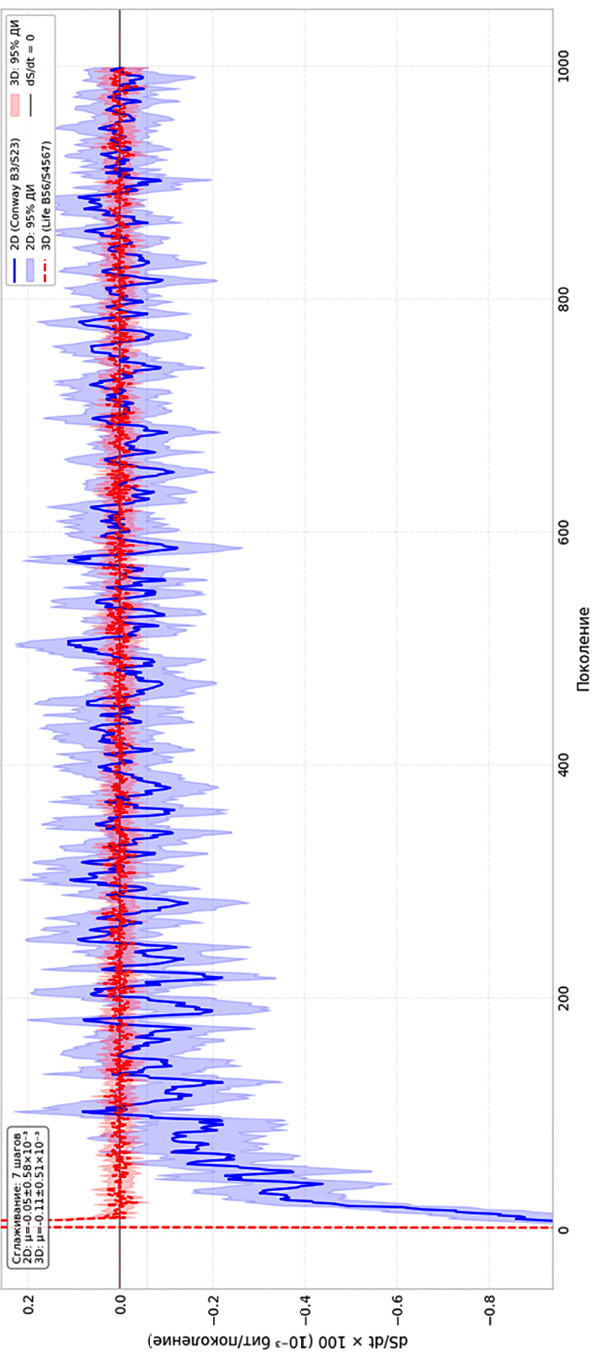
Рис. 3. Сравнение темпов изменения энтропии в 2D и 3D КА (сплошная – 2D, пунктирная линия – 3D) Примечание: составлен авторами по результатам данного исследования
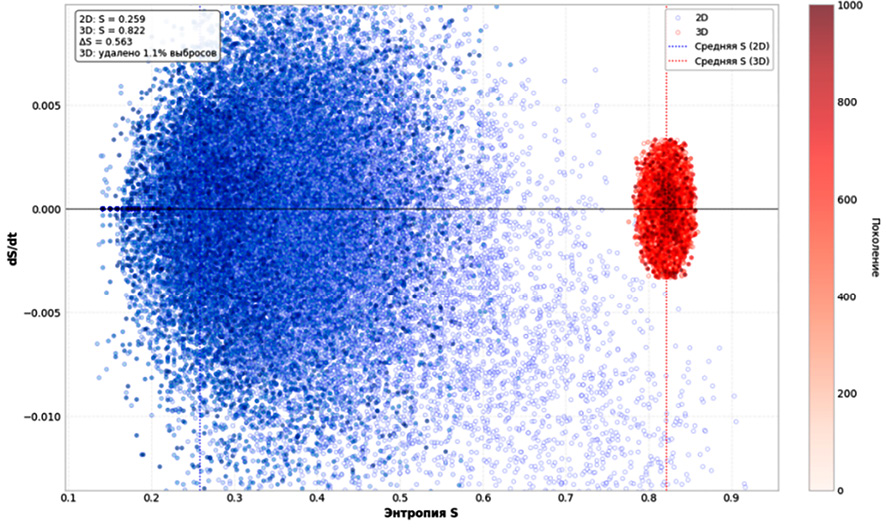
Рис. 4. Фазовый портрет динамики dS/dt от S для 2D и 3D КА Примечание: составлен авторами по результатам данного исследования
Таблица 2
Анализ ключевых метрик статистического анализа динамики 2D и 3D КА
|
Показатели |
2D |
3D |
|
Размер решетки |
50×50 = 2500 ячеек |
30³ = 27 000 ячеек |
|
Среднее число живых клеток (t = 1000) |
112 |
6 939 |
|
95 % ДИ для среднего |
[97, 127] |
[6892, 6986] |
|
SD числа живых клеток (t = 1000) |
55,7 |
169,8 |
|
CV числа живых клеток (t = 1000), % |
49,7 |
0,68 |
|
Начальная энтропия ⟨S(0)⟩, бит |
0,866 |
0,971 |
|
Финальная энтропия ⟨S(1000)⟩, бит |
0,255 |
0,822 |
|
95 % ДИ для финальной энтропии |
[0,230, 0,280] |
[0,819, 0,825] |
|
SD финальной энтропии |
0,090 |
0,011 |
|
CV(S) ансамбля (t = 1000), % |
35,3 |
1,3 |
|
CV временного ряда ⟨S⟩ (последние 200 шагов), % |
35,4 |
1,2 |
|
Характерное время релаксации τ, шаги |
~480 |
Не достигнуто |
|
Средняя скорость |dS/dt| (t = 0–200), ×10⁻³ бит/шаг |
12,9±1,2 (SD) |
25,6±0,8 (SD) |
|
Средняя скорость |dS/dt| (t = 800–1000), ×10⁻³ бит/шаг |
0,048±0,581 (SD) |
0,113±0,509 (SD) |
|
Амплитуда флуктуаций dS/dt (σ), ×10⁻³ бит/шаг |
1,71 |
2,18 |
|
Плотность кластеров (абс.)* |
5,34±0,74 (SD) |
20,56±0,14 (SD) |
|
Средний размер кластера, ед. кл. |
7,65±0,07 (SD) |
12,48±0,03 (SD) |
|
Доля живых клеток в кластерах, финал |
~38 % |
~67 % |
|
Корреляционная длина ξ, финал, ячейки |
1,18 |
1,63 |
Примечание: составлена авторами на основе полученных данных в ходе исследования.
Все величины вида «среднее ± интервал»: для числа живых клеток и энтропии указан 95 %-ный доверительный интервал (ДИ) для среднего по ансамблю; для скоростей и кластерных характеристик – стандартное отклонение (SD). CV числа живых клеток вычислен как SD/среднее; CV(S) для последних 200 шагов – это коэффициент вариации временного ряда средней по ансамблю энтропии ⟨S(t)⟩.
* Абсолютное количество связных кластеров на всю решетку: 2500 ячеек для 2D и 27 000 ячеек для 3D. Для сопоставления на 1000 ячеек плотность составляет ~2,14 (2D) и ~0,76 (3D).
Динамика, таким образом, отображает картину фрагментации активной фазы: вместо относительно связных областей мы видим переход к режиму разреженных и слабо взаимодействующих доменов.
В трехмерной модели динамика пространственной структуры выглядит иначе. Плотность кластеров здесь снижается умеренно – с 32,0 до 20,6, средний размер кластера практически не меняется (12,49 против 12,17 клетки), а корреляционная длина ξ сокращается незначительно: с 1,83 до 1,63 ячейки. Отношение финальных значений ξ₃ᴰ/ξ₂ᴰ ≈ 1,38 показывает, что в трехмерном случае пространственные корреляции оказываются более дальнодействующими; иными словами, объемный характер взаимодействий способствует сохранению целостности кластеров и поддерживает динамическую самоорганизацию, задаваемую правилами Life B56/S4567. Следовательно, в трехмерной среде формируются устойчивые домены активности, для которых характерны локальная самоподдержка и коллективная динамика, тогда как в двумерном варианте преобладает тенденция к структурной редукции и изоляции отдельных паттернов.
Доверительные интервалы (ДИ) для среднего числа живых клеток и средней энтропии построены двумя методами: bootstrap (5000 реплик, процентильный интервал) и нормальная аппроксимация (среднее ± 1,96 × SEM); оба подхода дали практически одинаковые результаты, и в табл. 2 приведены интервалы. Коэффициент вариации числа живых клеток вычислен как SD/среднее по ансамблю на финальном шаге, а CV(S) для последних 200 шагов представляет собой коэффициент вариации временного ряда усредненной по ансамблю энтропии ⟨S(t)⟩.
Сравнение финальных распределений числа живых клеток и энтропии между 2D и 3D по U-критерию Манна – Уитни дало p < 10⁻⁵ (N = 50). Доверительные интервалы в табл. 2 построены бутстрепом (5000 реплик, процентильный метод) и практически совпадают с нормальной аппроксимацией (среднее ± 1,96·SEM). При повторении всей серии из 50 прогонов с 10 разными стартовыми распределениями системы, средние значения числа клеток менялись не более чем на 2 % (2D) и 0,5 % (3D), а средней энтропии – не более чем на 1,5 % и 0,2 % соответственно.
Фазовый портрет в координатах dS/dt – S приведен на рис. 4. В двумерной системе после завершения переходного процесса траектория локализуется в компактной области с координатами S ≈ 0,259±0,025 бит и dS/dt ≈ 0. С точки зрения качественного анализа динамических систем такое поведение указывает на существование аттрактора – устойчивого макросостояния, к которому стремится ансамбль реализаций. Мы сознательно избегаем термина «точечный аттрактор» в строгом смысле, поскольку остаточные флуктуации энтропии свидетельствуют о сохранении стохастической компоненты. Тем не менее в пределах наблюдаемого временного окна система 2D демонстрирует выраженную сходимость к стационарному распределению с малым средним дрейфом, что оправдывает употребление понятия аттрактора в контексте статистической физики неравновесных систем. В трехмерной системе траектория локализована в узкой зоне со средним значением энтропии 0,822±0,003 бит, флуктуации производной минимальны (0,003), однако сходимости к точке не наблюдается. Разность средних энтропий между 3D и 2D равна 0,564 бит.
Далее на рис. 5 приведена фазовая траектория в координатах плотность – энтропия (ρ–S). Для двумерной модели: начальная точка (0,289; 0,866), конечная – (0,045; 0,255). Длина траектории L = 0,658. Потеря энтропии – 71 %. В трехмерной модели начальные значения (0,25; 0,971), конечные – (0,257; 0,822). Длина L = 0,868, отношение L₃ᴰ/L₂ᴰ = 1,31.
Несмотря на то, что траектория в трехмерном случае оказывается длиннее, это не свидетельствует о более выраженной эволюции макросостояния: на протяжении всего пути энтропия остается высокой (0,822), а плотность активных клеток меняется незначительно. Наблюдаемое увеличение траектории отражает скорее богатство микроскопических перестроек, которые не приводят к заметному упорядочению, но поддерживают высокую информационную насыщенность. Для двумерной модели за 480 шагов популяция снижается с 722 до 112±15 клеток (4,5 % площади решетки). Энтропия падает на 0,611 бит (71 % от исходной). Фазовые траектории выходят на точечный аттрактор (S = 0,255±0,025 бит). Коэффициент вариации составляет 49,7 %. Структура становится разреженной, преобладают детерминированные паттерны. В свою очередь, для трехмерной модели картина иная. Финальная численность – 6939±47 клеток (25,7 % объема). Энтропия снижается на 0,149 бит (15 %), финальное значение – 0,822±0,003 бит (82,2 % от максимума). Коэффициент вариации – 0,68 %. Система остается в квазистационарном режиме, выраженной тенденции к упорядочению не наблюдается.
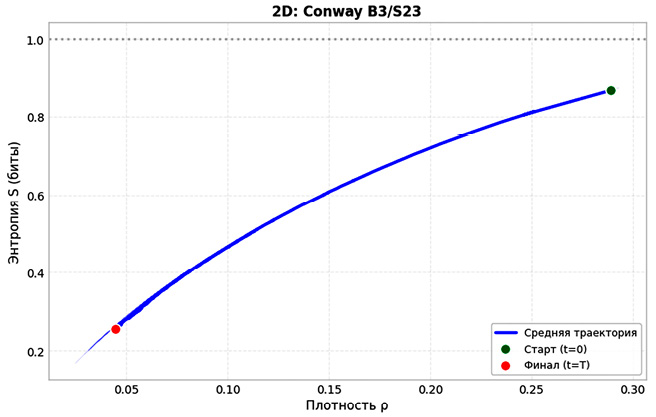
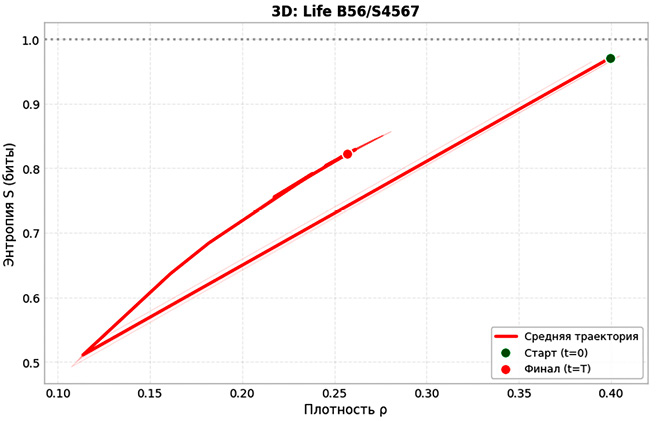
Рис. 5. Фазовая траектория в пространстве «плотность активных клеток – энтропия» (ρ−S) для 2D и 3D КА Примечание: составлен авторами по результатам данного исследования
Кластерный анализ дал следующие соотношения. В трехмерной модели средний размер кластера – 12,48 клетки, в двумерной – 7,65 (разница 63 %). Корреляционная длина в 3D составляет 1,63 ячейки, в 2D – 1,18 (на 38 % выше). Доля живых клеток, входящих в кластеры, в 3D – 67 %, в 2D – 38 %. Длина фазовой траектории в координатах (ρ–S) для 3D оказалась 0,868, а отношение L₃ᴰ/L₂ᴰ = 1,31. В исследованной конфигурации 3D-модель демонстрирует качественно иной тип динамики: вместо диссипативной релаксации, наблюдаемой в 2D, формируется метастабильная самоорганизация с устойчивыми объемными доменами, поддерживающими коллективную активность. Следует, однако, отметить, что данное различие может быть следствием совместного влияния размерности и режима обновления; для выделения чистого эффекта размерности необходимы дополнительные контрольные эксперименты.
Заключение
В работе проведено сравнение динамики двумерного и трехмерного стохастических клеточных автоматов при идентичных начальных условиях и усреднении по 50 независимым реализациям. Ключевой результат состоит в том, что в исследованных условиях размерность решетки оказалась сопряжена с качественным различием наблюдаемых динамических режимов. Например, 2D система демонстрирует диссипативное поведение – число активных ячеек падает с 722 до 112 (4,5 % решетки), энтропия снижается с 0,866 до 0,255 бит (потеря 71%), а фазовая траектория за ~480 шагов выходит на точечный аттрактор. При этом сохраняется высокая вариабельность остаточной активности (CV = 49,7 %). В 3D клеточном автомате, напротив, активность не затухает: финальная популяция составляет 6939 ячеек (25,7 % объема), энтропия остается близкой к максимальной (0,822 бит, потеря всего 15 %), а фазовые траектории после 1000 шагов не сходятся, совершая метастабильные блуждания. Статистическая устойчивость в трехмерном случае оказывается на порядок выше (CV = 0,68 %, что в десятки раз ниже, чем в 2D). Кластерный анализ дополняет картину, средний размер кластера в 3D составляет 12,48 против 7,65 клеток в 2D, корреляционная длина выше на 38 %, а доля живых клеток, входящих в кластеры, достигает 67 % против 38 % в 2D случае.
В проведенных опытах трехмерная модель демонстрирует метастабильную динамику с высокой энтропией, тогда как двумерная – быстрый переход к упорядоченному состоянию. Поскольку наряду с размерностью различались и протоколы обновления, однозначно приписать этот контраст повышению размерности нельзя; необходимы контрольные эксперименты с унификацией режима обновления. Тем не менее выявленные соотношения свидетельствуют о топологической обусловленности устойчивости неравновесных конфигураций. Дальнейший интерес представляет изучение репродуктивных свойств многомерных кластеров и их способности формировать функционально активные среды с длительным сохранением коллективной динамики.
Conflict of interest
Financing
Библиографическая ссылка
Клеммер П. С., Рави Кумар, Шебанов В. В. Исследование поведения стохастической энтропии в системе клеточного автомата 2D и 3D размерности // Международный журнал прикладных и фундаментальных исследований. 2026. № 2. С. 40-50;URL: https://applied-research.ru/en/article/view?id=13789 (дата обращения: 01.07.2026).
DOI: https://doi.org/10.17513/mjpfi.13789