
Scientific journal
International Journal of Applied and fundamental research
ISSN 1996-3955
ИФ РИНЦ = 0,556
CHANGE OF THE FUNCTIONAL STRUCTURE OF WOOD-DESTROYING FUNGI COMMUNITIES AS A REFLECTION OF FOREST STANDS STATE

Древоразрушающие базидиальные грибы являются неотъемлемой частью лесных экосистем разного генезиса, выполняя широкий спектр функций [8]. Однако если деятельность сапротрофных видов, обитающих на мертвой древесине и обеспечивающих возврат в круговорот экосистемы питательных веществ, можно оценить положительно с точки зрения сохранения и развития лесного фитоценоза, то в отношении фитопатогенных видов эта оценка не так однозначна. Грибы, обладающие фитопатогенными (биотрофными) свойствами, составляют важную группу, в состав которой входят виды, вызывающие гнилевое поражение деревьев, их ослабление и даже гибель. В природе роль этой группы грибов предопределена в качестве исполнителя функций структурной перестройки фитоценозов в их эволюционном развитии. В чистом виде, без вмешательства прочих биотических и абиотических факторов, грибы этой группы должны обеспечивать постоянный в определенных размерах отпад части деревьев из древостоя, объем которого, выраженный в фитомассе, должен быть соизмерим с ее приростом для конкретного положения биогеоценоза в сукцессионном ряду его развития [14].
Соответственно, с хозяйственной и научной точек зрения, роль дереворазрушающих грибов может быть оценена двояко: с одной стороны, дереворазрушающие грибы повреждают вегетирующие деревья, иногда приводя к выпадению из древостоя значительного количества экземпляров деревьев, следствием чего становится изреживание насаждений и изменение их микроклимата; с другой стороны, фитопатогенные виды грибов вызывают гнили именно ослабленных деревьев, приводя к отпаду из древостоя наименее устойчивых деревьев [16].
Именно фитопатогенные грибы наиболее эффективно выполняют регуляторную функцию в лесных экосистемах [11]. Выполнение всеми компонентами экосистемы регуляторных функций обеспечивает ее саморегуляцию, которая дает ей возможность при различных неблагоприятных условиях, вызванных, в частности, антропогенной деятельностью, регулировать интенсивность обмена с окружающей средой с целью поддержания гомеостаза. Такая саморегуляция может осуществляться в результате адаптивных перестроек структуры связей между отдельными элементами системы, направленных на повышение устойчивости экосистемы в новых условиях [5].
Регуляция лесной экосистемы дереворазрушающими грибами производится несколькими способами: посредством прямой регуляции древостоев фитопатогенными грибами; через изменение количества поступления питательных веществ в почву за счет изменения интенсивности процессов деструкции [11]; через регуляцию численности других групп гетеротрофных организмов, связанных с ксилотрофными грибами трофическими связями (в первую очередь это касается насекомых – мицетофагов [4, 6].
В.А. Мухин [7] предложил различать два основных пути разложения древесины – сапротрофный и фитопатогенный; наиболее существенным различие между ними заключается в том, что вследствие деятельности ксилотрофных грибов, заселяющих вегетирующие древесные растения, уменьшается продолжительность жизни деревьев и, тем самым, ускоряется поступление веществ древесины в «цепи разложения» лесных биогеоценозов. То есть фитопатогенный путь ведет к интенсификации круговорота веществ в лесных экосистемах.
Таким образом, грибы оптимизируют лесную экосистему, способствуя формированию древостоев, более соответствующих данному типу экотопа [13].
При этом возникает вопрос: какие перестройки претерпевает структура грибного сообщества (в первую очередь – функциональная) в ходе трансформации древостоев леса до субклимаксового состояния? И есть ли возможность, анализируя структуру микоценоза, сделать вывод о стадии развития лесной экосистемы?
Для ответа на эти вопросы нами был проведен анализ структуры сообществ базидиальных дереворазрушающих грибов ряда искусственных насаждений в Южном Приуралье (Оренбургская область).
Материалы и методы исследования
Для оценки структурной динамики сообществ дереворазрушающих грибов нами осуществлялись экспедиционные исследования в 1994-2013 гг., охватывавшие разные типы лесов в разных районах Оренбургской области. Значительная часть исследований проводилась в искусственных насаждениях разного породного состава, так как эти насаждения преимущественно монодоминнатные и одновозрастные, что позволяет нивелировать значение лесовосстановительных процессов на формирование и развитие грибных сообществ. Древостои искусственного генезиса в структурном отношении далеки от оптимального строения и не обладают эволюционно выработанными механизмами устойчивости к дестабилизирующему воздействию внешних факторов. Такие механизмы могут быть выработаны только при прохождении этих искусственных экосистем через серию сукцессионных перестроек, в ходе которых будет увеличиваться соответствие структурно-функциональных характеристик экосистемы конкретным условиям биотопа [11, 13]. При этом грибы являются, разумеется, не единственным фактором, определяющим динамику структур сообществ – к микогенному влиянию присоединяются изменения гидрологического режима, воздействия ветров, антропогенная деятельность в разных формах и т.д.
Объектом исследований являлись базидиальные ксилотрофные макромицеты, преимущественно афиллофороидные грибы, являющиеся основными возбудителями стволовых и корневых гнилей древесных растений, а также производящие деструкцию детрита в исследуемых лесах региона.
Сбор образцов производился методом маршрутного сбора. На маршруте проводилось описание биотопов и субстрата, на котором обитали грибы; оценка численности ксилотрофных базидиомицетов основывалась на определении в 2-метровой полосе учета количества древесных остатков, на которых развивается тот или иной вид [7].
В общей сложности было собрано и определено более 1200 образцов плодовых тел дереворазрушающих макромицетов. Идентификация собранных образцов была произведена с использованием русскоязычной и зарубежной определительной литературы [1-3, 15, 17, 18, 20].
При анализе структуры грибных сообществ применялись подходы, разработанные В.А. Мухиным [7], М.А. Сафоновым [11]. Для определения структурных особенностей использовался Индекс Бергера-Паркера, как показатель доминирования преобладающего вида.
Результаты исследования и их обсуждение
В результате исследований было установлено, что в состав фитопатогенного комплекса грибов искусственных насаждений Южного Приуралья входят 22 вида, из которых половина является факультативными фитопатогенами, т.е. чаще всего были обнаружены на сухостойных деревьях и разных фракциях валежа [12].
Прочие виды более-менее постоянно встречаются на живых деревьях. Особенно широкое распространение в искусственных насаждениях получили такие виды, как трутовик настоящий (Fomes fomentarius (L.: Fr.) Fr.), трутовик чешуйчатый (Polyporus squamosus Huds.:Fr.), сливовый трутовик (Phellinus tuberculosus (Baumg.) Niemela), опенок зимний (Flammulina velutipes (Curt.: Fr.) Sing.), серно-желтый трутовик (Laetiporus sulphureus (Bull.: Fr.) Murrill). Эти виды являются активными фитопатогенами как в искусственных, так и в естественных насаждениях региона [10, 11].
Наиболее активным и наиболее распространенным фитопатогенном является трутовик настоящий (Fomes fomentarius). Южном Приуралье этот вид отмечен на древесине всех родов древесных растений, кроме сосны, лиственницы и жимолости [10]. Особенно велика значимость гриба в микоценозах березняков и осинников, где с его участием происходит деструкция валежной древесины, пней, сухостоя и вегетирующих деревьев. Максимальная зараженность отмечена в перестойных березняках и осинниках (до 30 %) [11].
Наиболее широко группа фитопатогенных грибов представлена на яблонях, немного меньше видов было отмечено на березе и клене (рис. 1).
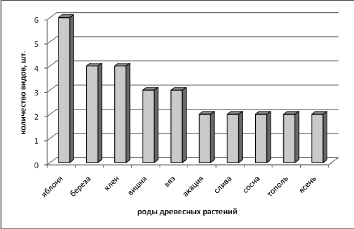
Рис. 1. Численность видов дереворазрушающих грибов, отмеченных в искуственных насаждениях на вегетирующих деревьях разных родов в районе исследований
По относительному и абсолютному участию фитопатогенных видов сообщества ксилотрофных грибов обследованных искусственных насаждений заметно отличались (таблица).
Варьирование значения биотрофных видов грибов в сообществах в искусственных насаждениях разных типов
|
Типы насаждений |
Участие биотрофных видов |
||
|
Индекс Бергера-Паркера |
доля видов, % |
относительная численность видов, % |
|
|
Монодоминантные насаждения |
|||
|
березовые посадки в целом |
0,39 |
9,8 |
20,0 |
|
широколиственные насаждения |
0,38 |
22,8 |
35,6 |
|
сосновые насаждения |
0,24 |
14,3 |
9,5 |
|
Полидоминантные насаждения |
|||
|
Широколиственные насаждения |
0,30 |
18,8 |
23,5 |
|
Средние значения по группам насаждений |
|||
|
Среднее арифметическое |
0,36 |
16,1 |
23,8 |
|
Lim |
0,15-0,66 |
5,0 – 17,6 |
7,5-37,2 |
Особенно заметно варьирует относительная численность видов-фитопатогенов, т.е. доля плодовых тел этих видов от общего количества базидиом. отмеченных в данном биотопе. Для сообществ, в которых доминирующее положение занимали фитопатогены, мы провели анализ по группам насаждений. Анализ показал, что по показателям индекса Бергера-Паркера, позволяющего оценить степень доминирования преобладающего вида, микоценозы насаждений разных типов мало отличались: минимальный показатель был получен для микоценозов сосновых насаждений (0,24).
Доля фитопатогенных видов в сообществах варьировала заметнее. Более разнообразно биотрофы были представлены в широколиственных насаждениях. Наименьшее разнообразие было отмечено в березовых насаждениях; в этих сообществах основным видом-фитопатогеном являлся трутовик настоящий (Fomes fomentarius (L.: Fr.) Fr.).
По относительной численности плодовых тел фитопатогенных видов также выделяются микоценозы широколиственных насаждений, а минимальное участие было характерно для сообществ грибов сосновых насаждений.
Анализ функциональной структуры изученных микоценозов показал, что во всех сообществах доминирующее положение занимали 1-2 вида, встречи остальных видов были малочисленными или имели случайный характер. В большинстве случаев картина распределения частот встречаемости видов была достаточно однотипна. Однако следует отметить, что в тех сообществах, где доминирующую позицию занимали фитопатогенные виды, линии трендов распределения видов по численности аппроксимировались степенными уравнениями, а в прочих (т.е. с доминированием сапротрофных видов) – логарифмическими уравнениями (рис. 2).
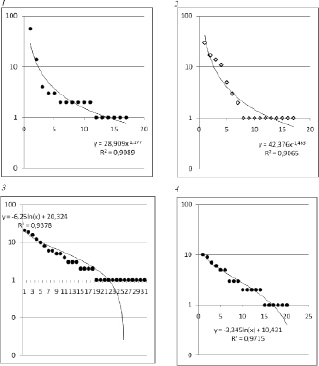
Рис. 2. Функциональная структура некоторых изученных сообществ ксилотрофных грибов. 1,2 – сообщества с доминированием фитопатогенного вида (Fomes fomentarius), 3,4 – сообщества с доминированием сапротрофных видов
Как известно, кривые значимости видов [19] считаются наилучшим способом графического представления двух компонентов разнообразия – видового богатства и выравненности видов [9]. При этом большинство получаемых графиков можно свести к 3 вариантам, каждый из которых представляет определенные условия дифференциации экологических ниш в сообществе. Исходя из этой классификации, сообщества грибов, в которых доминируют фитопатогенные виды, могут быть отнесены к I типу, т.е. представляет собой геометрический ряд с приоритетами в захвате ниш, где каждый вид занимает в первую очередь свободную нишу, не перекрывающуюся другими; такие закономерности характерны для некоторых растительных сообществ в экстремальных условиях [9]. В грибных сообществах с доминированием сапротрофных видов ниши многомерны и перекрываются, что обуславливает логнормальное распределение видов по значимости [21]. Эта закономерность отражает наиболее вероятную, широко распространенную модель, особенно для ненарушенных сообществ [9].
Следовательно, функциональная структура сообществ дереворазрушающих грибов развивается в направлении снижения доли фитопатогенных видов и увеличения участия сапротрофов, что соответствует более устойчивому состоянию лесных экосистем.
Следует отметить, что искусственные насаждения обладают благоприятными условиями для активного развития фитопатогенного пути, так как их древостои состоят из деревьев одного возраста, подвергающихся более-менее длительное время одним и тем же неблагоприятным воздействиям. Соответственно, появление условий для проникновения и развития грибной инфекции являются значимыми также для всех особей популяции, т.е. создаются условия для активного распространения гнилей. Вероятно, именно по этой причине появившиеся в искусственных насаждениях фитопатогенные грибы начинают активно наращивать свою численность, выходят на доминирующую позицию и остаются на ней до осуществления окончательной структурной перестройки древостоя, т.е. отпада из него ослабленных деревьев.
Таким образом, фитопатогенное состояние изученных искусственных насаждений нельзя охарактеризовать однозначно. В тех насаждениях, где присутствует большое количество старых экземпляров деревьев или деревьев, имеющих механические повреждения вследствие действия неблагоприятных климатических факторов или антропогенных воздействий, биотрофные виды активно участвуют в регулировании структуры древостоя. В старовозрастных насаждениях с высоким видовым разнообразием дереворазрушающих грибов – сапротрофов структура древостоев не нуждается в подобной корректировке и подобные микоценозы могут существовать неопределенно долго. Многие изученные микоценозы искусственных насаждений включают один-два вида – фитопатогена. Фитопатогенное состояние этих искусственных насаждений можно охарактеризовать как хорошее. Следовательно, среди искусственных насаждений области наблюдается широкая дисперсия по уровню фитопатогенного поражения. Однако в подавляющем большинстве случаев распространение фитопатогенных видов не носит характер эпифитотии.
Заключение
Проведенный анализ позволяет придти к выводу, что изменение структуры древостоев не только сопровождаются изменением сообществ дереворазрушающих грибов, но и, в определенной степени, определяются этими изменениями. Регуляторная функция ксилотрофных грибов наиболее ярко реализуется за счет деятельности фитопатогенных видов, которые обеспечивают отпад из древостоя поврежденных или перестойных деревьев, оптимизируя лесной фитоценоз. По мере осуществления этих структурных перестроек, меняется и микоценоз: снижается доля фитопатогенов, увеличивается видовое разнообразие грибов, за счет сапротрофных видов. В итоге формируется функциональная структура сообщества, ориентированная на более эффективное использование ресурсов среды.
Соответственно, структурные особенности (в особенности – функциональная структура) грибного сообщества являются отражением состояния древостоя и анализ структуры микоценозов позволяет оценить стадию формирования древостоя.
Библиографическая ссылка
Сафонов М.А., Маленкова А.С. ИЗМЕНЕНИЕ ФУНКЦИОНАЛЬНОЙ СТРУКТУРЫ СООБЩЕСТВ ДЕРЕВОРАЗРУШАЮЩИХ КАК ОТРАЖЕНИЕ СОСТОЯНИЯ ДРЕВОСТОЕВ // Международный журнал прикладных и фундаментальных исследований. 2014. № 8-1. С. 72-77;URL: https://applied-research.ru/en/article/view?id=5642 (дата обращения: 25.05.2026).