
Scientific journal
International Journal of Applied and fundamental research
ISSN 1996-3955
ИФ РИНЦ = 0,556
MIGRATIONS OF HARP SEALS OF WHITE SEA POPULATION AT THE FIRST YEAR OF LIFE ACCORDING TO DATA OF SATELLITE TELEMETRY

Гренландский тюлень Phoca groenlandica является наиболее многочисленным видом ластоногих в Северной Атлантике. В пределах обширного ареала вид образует три обособленные популяции, названные по основным местам их размножения: ньюфаундлендская (включая залив Св. Лаврентия), ян-майенская и беломорская [4]. Общая численность вида в настоящее время оценивается в 8,5–9 млн особей.
Сезонные миграции беломорской популяции гренландского тюленя охватывают значительную часть Северной Атлантики с прилегающими морями: Баренцевым, Гренландским и Норвежским, а также Карское море. Миграция идет в направлении от северной границы Белого моря на северо-запад вдоль берегов Скандинавского полуострова. Тюлени достигают вод у архипелага Шпицберген, затем следуют вдоль ледовой кромки между архипелагами Шпицберген и Земля Франца-Иосифа, далее движутся на юг вдоль западного побережья архипелага Новая Земля на юго-восток Баренцева моря и затем – в Белое море [5, 8, 9].
В период летних миграций тюлени обитают в Баренцевом море и других районах Северной Атлантики, встречаются в Карском море [2, 5, 7]. Зимой они заходят в Белое море для спаривания и размножения, где с декабря по май ведут ледовый образ жизни, образуя массовые залежки. Льды с детными (щенными) залежками постепенно выносятся из центральных районов Белого моря на северо-восток и далее в Баренцево море.
Детеныши гренландского тюленя находятся в Белом море с февраля по май. Первые этапы их постнатального развития (молочное кормление, первая линька, переход на самостоятельное питание) проходят на дрейфующем льду. В это время они адаптируются к обитанию в воде. Сроки щенения гренландского тюленя в Белом море длительное время, начиная с 1925 г., остаются неизменными [3]. В то же время, в связи с оскудением кормовой базы (истощение запасов сайки и мойвы, основных объектов питания), а также колебаниями ледовых условий в Белом море в последние годы пути миграций гренландских тюленей к основному месту нагула, кромке льдов Баренцева моря, могут существенно изменяться.
К.К. Чапский [5] не исключал возможность миграции гренландских тюленей старше 1 года вдоль ледовой кромки на северо-восток Баренцева моря, затем – к о. Колгуев и далее вдоль ледовой кромки до Гусиной Земли, а также на запад вдоль побережья Мурмана. Согласно схеме, представленной Е. Сивертсеном [10], приплод гренландского тюленя при выходе из Белого моря в апреле мигрирует в северном и северо-восточном направлениях. В более поздних литературных источниках «восточная» миграция гренландского тюленя полностью исключалась [6]. Тем не менее, материалы, полученные в результате спутникового мечения половозрелых гренландских тюленей в Белом море в 1995–1996 гг., впервые проведенного В.А. Потеловым и соавторами [2], показали, что часть животных в мае – июне мигрирует и в восточном направлении – в Печорское море.
Точных данных о миграциях детенышей гренландских тюленей беломорской популяции в различные периоды года нет. Цель настоящей работы – определить направления и сроки перемещения детенышей гренландского тюленя в Белом и Баренцевом морях в первые месяцы их жизни.
Материалы и методы исследования
Для проведения работ по мечению тюленей были использованы датчики спутниковой телеметрии (ДСТ) «Пульсар» производства ЗАО «ЭС-ПАС» (Россия). ДСТ функционируют в составе спутниковой системы Argos, позволяющей определять местоположение животного с точностью от 350 до 1500 метров. После установки радиомаяка на животное и приведения в действие его передатчика, он начинает излучать импульсы мощностью 500 мВт на частоте 401,650 мГц ± 30,0 кГц каждые 50 секунд. Продолжительность работы радиомаяка зависит от числа передач сообщений и определяется поведением животного, поскольку передача осуществляется, когда спина животного с закрепленным на ней ДСТ находится вне воды, и электрическая цепь, образованная морской водой и электродами на верхней стороне датчика, разрывается.
Отлов тюленей производили по разрешению Баренцево-Беломорского управления Федерального агентства по рыболовству. Научная группа высадилась на лед с вертолета (МИ-8) в точке с координатами 64 °42’23’’ с.ш./38 °56’02’’ в.д. на границе горла и бассейна Белого в 20–25 км от берега. Тюлени были отловлены после завершения молочного вскармливания и начала первой, ювенильной линьки (переход из стадии «белек» в стадию «серка»). Для завершения линьки был выбран безлюдный район на мысе Кяндский в Онежском заливе Белого моря, где щенки были помещены в вольер 5×10 м из металлической сетки-рабицы с пластиковым покрытием.
6 апреля, когда все тюлени лишились эмбрионального меха, установили ДСТ (рис. 1). Для установки датчиков использовали двухкомпонентный эпоксидный клей, а сами они были заранее приклеены к подложке из сети с ячеей 12 мм. Было проведено тестирование ДСТ, получен ряд откликов через сеть Argos. В системе Argos датчикам присвоены идентификационные индексы: 84578; 97601; 97602; 97603.
Результаты исследования и их обсуждение
8 апреля 4 меченых тюленя (1 самец и 3 самки) были выпущены на лед в точке с координатами 66 °10’17’’ с.ш./40 °10’11’’ в.д. В этом районе наблюдали скопление серок гренландского тюленя. Первые данные о местоположении меченых животных на льдах в Горле Белого моря были получены 9 апреля (рис. 2).
Миграции тюленей в Белом море
В первую неделю тюлени дрейфовали на льдинах в направлении о. Моржовец. В это время льды находились в зоне действия Беломорского течения, которое начинается от м. Зимнегорский, следует вдоль Зимнего берега до м. Инцы, на север к о. Сосновец и далее – в Баренцево море [1]. В середине второй декады апреля тюлени стали сходить в воду и перемещаться по акватории моря, попадая под воздействие течений.

Рис. 1. Щенок гренландского тюленя с датчиком спутниковой телеметрии на льду Белого моря
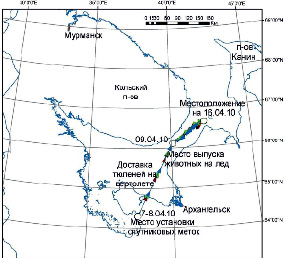
Рис. 2. Перемещения тюленей с метками № 84578, 97601, 97602 и 97603 c 7 по 16 апреля 2010 г. по данным спутниковой телеметрии Argos
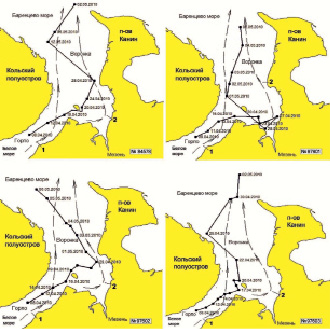
Рис. 3. Схема миграции гренландских тюленей с ДСТ №№ 84578, 97601, 97602 и 97603 в Белом море. Обозначения: 1 – Беломорское течение, 2 – Мезенское течение
В Белом море все серки гренландского тюленя с установленными ДСТ мигрировали в северном направлении в течение апреля и начале мая. Тюлени придерживались основных течений – Мезенского и Беломорского. Однако серка № 84578 на короткое время вновь вернулась в Белое море. Несмотря на разные маршруты, к 6 мая все тюлени покинули Белое море.
Серка № 84578 в конце апреля достигла м. Конушин благодаря Мезенскому течению. В дальнейшем тюлень двигался на север, и в начале мая достиг Баренцева моря. Во второй декаде мая он вновь вернулся в Белое море (рис. 3).
Серка № 97601 с середины второй декады апреля по конец апреля находилась в кутовой части Мезенского залива. Значительные приливно-отливные течения, характерные для этого района, опасны для молодых тюленей в период сжатия льдов. Уменьшение числа сигналов в этот период времени позволяет предположить, что серка активно двигалась по акватории, используя стационарную полынью в южной части залива и большую часть времени находилась в воде. В конце апреля этот тюлень мигрировал на северо-запад и, достигнув Беломорского течения, продолжил двигаться на север, а к середине первой декады мая достиг границы Белого моря (рис. 3).
Серка № 97602 практически полностью повторила путь серки № 84578, отстав от нее на 2-3 дня, но этот тюлень не возвращался обратно в Белое море (рис. 3).
Серка № 97603 оказалась самой быстрой, вероятно, потому, что не выходила из Беломорского течения. К 22 апреля этот тюлень уже находился у м. Конушин, а 30 апреля вышел в воды Баренцева моря (рис. 3).
Полученные результаты позволяют сделать следующие заключения. Очевидно, что серки покидают Белое море в апреле-начале мая, дрейфуя вместе со льдами. В Горле и Воронке моря большое значение для миграции гренландских тюленей имеют выносные течения, главным из которых является Беломорское. В 2010 г. тюлени мигрировали из Белого моря в течение 3–4 недель, покинув его в начале мая. При этом они использовали стационарные полыньи в Мезенском заливе и Воронке, где задерживались на некоторое время.
Миграции тюленей в Баренцевом море
Миграционные маршруты молодых гренландских тюленей с ДСТ в Баренцевом море отличались большим разнообразием. Тюлени распределялись по всей его акватории в течение длительного нагульного периода. Наиболее предпочтительными местами обитания для тюленей оказались районы кромки многолетних дрейфующих льдов между о-вами Шпицберген и Земля Франца-Иосифа. Наблюдения показали, что молодые тюлени могут достигать западного побережья о. Шпицберген, где, возможно, смешиваются с тюленями ян-майенской популяции. В то же время серки длительное время могли держаться и вдали от кромки льдов, в пелагической части Баренцева моря.
Серка №84578 в третьей декаде мая мигрировала по «восточному» направлению, используя Канинское течение, к северным склонам Канино-колгуевского мелководья и Северо-Канинской банки (рис. 4). В начале июня тюлень ушел в район Гусиной банки, где оставался более 1 месяца. Дальнейшая миграция на север началась в конце 1-й декады июля, а уже в конце июля тюлень пересек 74 ° с.ш. В третьей декаде августа тюлень достиг ледовой кромки между островами Земля Франца-Иосифа и Шпицберген, где и оставался до середины сентября, пока метка не прекратила работу.
Серка № 97601 мигрировала по «западному» направлению в районе Мурманского мелководья, но затем тюлень двинулся на восток от 40 ° в.д. (рис. 4). В июне и июле тюлень держался в районе 74 ° с.ш. между 32 ° и 48 ° в.д., в районе Центральной возвышенности. В начале августа тюлень активно перемещался почти до 54 ° в.д. и вновь вернулся в исходный район в середине августа. В конце августа тюлень пересек 76 ° с.ш. и, двигаясь на северо-восток, к концу первой декады сентября достиг западной части Возвышенности Персея. Не доходя до ледовой кромки, тюлень продолжил миграцию на восток, достигнув к концу сентября район м. Желаний (о. Новая Земля). Затем тюлень начал двигаться в южном направлении и в середине третьей декады ноября уже находился на припайных льдах о. Новая Земля в районе пролива Маточкин Шар (м. Сухой Нос). В первых числах декабря 2010 г. тюлень достиг однолетних льдов между о. Колгуев и южной оконечностью о. Новая Земля, где и оставался до начала февраля 2011 г.
Тюлень №97602 покинул акваторию Белого моря в середине 1-й декады мая и быстро двигался на запад вдоль Кольского побережья, придерживаясь «западной» ветви миграции. Однако уже в середине мая он изменил направление на север и северо-восток, достигнув в начале июня района Западного склона Гусиной банки, расположенного в зоне теплых течений. Только почти через месяц тюлень ушел из этого района на северо-восток. В начале 2-й декады июля тюлень поднялся выше 76 ° с.ш. в воды о. Земля Франца-Иосифа, но так и не достиг ледовой кромки. По какой-то причине тюлень не стал подниматься в зону льдов, а в начале сентября пошел на юго-восток, в направлении к о. Новая Земля, и в середине сентября вернулся в район Гусиной банки. Затем серка продолжила миграцию на северо-восток к Земле Франца-Иосифа. В начале второй декады октября тюлень достиг ледовой кромки и оставался там до потери связи с ДСТ 04.11.2010 г.
Миграционный путь тюленя с датчиком № 97603 до 31.12.2010 оказался самым продолжительным. Этот тюлень покинул Белое море раньше остальных – в конце апреля и, двигаясь сначала на северо-восток, а потом на северо-запад, достиг ледовой кромки к началу ноября. Тюлень оставался в открытой пелагической части моря у ледовой кромки между о-вами Шпицберген и Земля Франца-Иосифа до конца 2010 г. (рис. 5). В начале января 2011 г. тюлень мигрировал вдоль ледовой кромки в западном направлении, но уже в середине января переместился значительно южнее, к о. Медвежий, а затем двигался в восточном направлении, придерживаясь ледовой кромки. Достигнув в середине апреля самой северо-восточной точки своего пути, тюлень повернул на юг вдоль припайных льдов о.Новая Земля. К 28 апреля он достиг юго-восточной части Баренцева моря (район Канинско-Колгуевского мелководья), где и оставался на льдах до 7 мая. У гренландских тюленей в данный период происходит линька, и ДСТ, очевидно, был потерян.
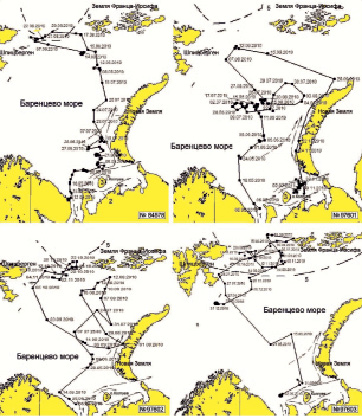
Рис. 4. Схема миграции гренландских тюленей с ДСТ №№ 84578, 97601, 97602 и 97603 в Баренцевом море. Обозначения: 1 – Беломорское, 2 – Канинское, 3 – Колгуевско-Печорское, 4 – Новоземельское течения, соответственно, 5 – кромка дрейфующих льдов на 01.08.2010
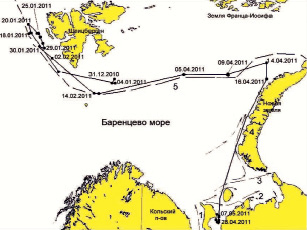
Рис. 5. Схема миграции гренландского тюленя № 97603 в Баренцевом море в 2011году. Обозначения те же, что на рис. 4. Граница ледовой кромки указана на 01.02.2011
В Баренцевом море в мае дрейфующие льды с залежками попадают в Канинское течение, благодаря которому тюлени совершают миграцию на север через восточную часть Баренцева моря. Летом тюлени следуют по Новоземельскому течению, задерживаясь на значительный срок в районах Гусиной банки, Возвышенности Персея и др. Это так называемое «восточное» направление миграции. Миграция сеголетков на север от скандинавского побережья Баренцева моря в 2010 г. не наблюдалась, хотя два тюленя до середины мая и двигались в западном направлении, но затем достаточно быстро вернулись в зону действия Новоземельского течения. Следует отметить, что «западное» направление ранее указывалось в качестве основного пути миграции для гренландских тюленей беломорской популяции [8, 9].
Заключение
Появление тюленей у ледовой кромки на севере Баренцева моря в 2010 г. было весьма растянуто по времени – с конца августа до начала ноября. Два тюленя, достигнув кромки льдов, периодически уходили от неё на большое расстояние в открытое море. Третий тюлень оставался на севере у ледовой кромки в течение всего периода наблюдения и даже совершил миграцию вдоль ледовой кромки в воды Норвежского моря у западного Шпицбергена (№ 97603). Аналогичное распределение тюленей в северной части Баренцева моря было получено ранее в результате мечения взрослых гренландских тюленей [9]. Однако наши наблюдения показали, что не все сеголетки во время летней миграции достигают северной кромки льдов, один из четырех тюленей с ДСТ держался в открытой части моря вплоть до образования припайных льдов у о-вов Новая Земля и на юго-востоке (№ 97601). Присутствие гренландских тюленей на льдах в юго-восточной части Баренцева моря в ноябре–феврале было неоднократно описано для тюленей разных возрастных групп [9]. Как показано в нашем исследовании, молодые тюлени (сеголетки) приходят в эти районы одновременно с тюленями других возрастных групп (1 год и старше).
Также следует отметить, что два из четырех меченых тюленей во время обратной миграции зимой вернулись на юго-восток Баренцева моря (Чешская губа и о. Колгуев), но не зашли в Белое море (метки остальных двух тюленей перестали работать в ноябре и декабре 2010 г.). Тем не менее, можно утверждать, что однолетние льды Чешской губы и прилегающие районы на юго-востоке Баренцева моря (Канин-Колгуев-Новая Земля) являются местом линьки молодых гренландских тюленей.
Таким образом, установка датчиков спутниковой телеметрии, впервые осуществленная на детенышах гренландского тюленя беломорской популяции, позволяет получать значительные массивы информации о сроках и путях миграции тюленей в первые месяцы жизни в зависимости от климатических и экологических условий. Дальнейший детальный анализ материалов спутниковой телеметрии в сопоставлении с данными о гидрометеорологической обстановке и наличии пищевых ресурсов в районах миграций внесет вклад в понимание тенденций развития популяций данного вида морских млекопитающих, играющего существенную роль в экосистеме Баренцева моря.
Библиографическая ссылка
Светочев В.Н., Светочева О.Н., Кавцевич Н.Н. МИГРАЦИИИ ГРЕНЛАНДСКИХ ТЮЛЕНЕЙ БЕЛОМОРСКОЙ ПОПУЛЯЦИИ НА ПЕРВОМ ГОДУ ЖИЗНИ ПО ДАННЫМ СПУТНИКОВОЙ ТЕЛЕМЕТРИИ // Международный журнал прикладных и фундаментальных исследований. 2015. № 12-3. С. 485-491;URL: https://applied-research.ru/en/article/view?id=7961 (дата обращения: 02.08.2026).