
Scientific journal
International Journal of Applied and fundamental research
ISSN 1996-3955
ИФ РИНЦ = 0,556
ADAPTIVE REACTIONS OF THE RESPIRATIVE PART OF LUNG UNDER LONGTERM HYPOXIC HYPOXY

Одним из важных факторов внешней среды является гипоксия, которая оказывает воздействие на все системы организма, включая органы дыхания. Изменения в легких прежде всего проявляются в состоянии легочного сурфактанта и ультраструктуре аэрогематического барьера. В настоящее время пристальное внимание уделяется адаптации к условиям высокогорья, экспериментальным аналогом которого является нормобарическая многократная гипоксия, развающаяся при вдыхании газовой смеси с содержанием кислорода 10–14 %. Как известно, нормобарическая гипоксия характеризуется снижением парциального давления кислорода в крови, вызывая развитие адаптивных, приспособительных реакций, направленных на уменьшение кислородной недостаточности. Литературные данные свидетельствуют в том, морфологические изменения при адаптации к гипоксии носят самый разнообразный характер, зависящий от мощности и времени воздействия раздражающего фактора [1–3]. По данным работы [4] при многократной гипоксической нагрузке 10 ежедневными 30-минутными сеансами у крыс линии Вистер наблюдались адаптивные изменения стрессорной системы. При этом установлено, что долговременные адаптивные реакции легких при гипоксической нагрузке реализуются повышением функциональной активности и числа эффекторных клеток: макрофагов, тучных клеток, лимфоцитов и нейтрофилов. Однако морфологическая оценка компенсаторно-приспособительных реакции не включала изучение респираторного отдела легких на ультраструктурном уровне. Поэтому нами была поставлена цель изучить адаптивные ултраструктурные реакции аэрогематического барьера легких при долговременной гипоксической гипоксии.
Эксперименты были проведены на половозрелых беспородных крысах-самцах массой 140–160 г. Первая группа животных была контрольной. Вторая подвергалась воздействию гипоксической гипоксии путем ежедневного вдыхания газовой смеси воздуха и азота (1:1) с содержанием кислорода 12 % в камере в течение 30 мин на протяжении 20 дней согласно методике [5].
Для электонно-микроскопического исследования кусочки легких фиксировали в 2,5 % растворе глютаральдегида с постфиксацией в 1 % растворе четырехокиси осмия, проводили по общепринятой методике и заключали в эпон. Ультратонкие срезы контрастировали уранилацетатом цитратом свинца по Рейнольдсу. Исследования проведены на электронном микрокопе ЭМВ-100Л.
Ультраструктурные изменения в респираторном отделе крыс при гипоксической гипоксии. Альвеолоциты 1-го типа характеризовались наличием крупного ядра, занимающего почти весь обьем цитоплазмы. Контуры ядерной оболочки волнистые, содержание эухроматина высокое, распределение гетерохроматина, примаргинальное. Цитоплазма высокой электронной плотности вследствие обилия свободных рибосом и полисом содержала гиперплазированные канальцы гранулярного эндоплазматического ретикулума, богатые мембраносвязанными рибосомами и содержащие в просвете хлопьвидный материал умеренной электронной плотности. Отдельные канальцы гранулярного эндоплазматического ретикулума были слегка расширены. Митохондрии в основным крупные. В митохондриях различимы матрикс умеренной плотности и четко выраженные кристы. Отдельные митохондрии имели набухший вид. Аппарат Гольджи развит хорошо. Вдоль апикальной поверхности плазмалеммы располагались мелкие пиноцитозные пузырьки, форма устьев которых свидетельствовала об их перемещении в сторону интерстиция. Базальная поверхность плазмалеммы была неровной. Базальная мембрана выглядела разрыхленной. Уплощенные периферические отделы альвеолоцитов 1-го типа имели высокую электронную плотность и неравномерную толщину. В утолщенных отделах цитоплазмы располагались крупные митохондрии, канальцы ретикулума, многочисленные свободные рибосомы. Изредка в цитоплазме альвеолоцитов 1-го типа наблюдались мелкие осмиофильные тельца и гранулы разрушенного сурфактанта (рис. 1). Апикальная поверхность формировала многочисленные длинные и тонкие цитоплазматические выросты, способствующие формированию крупных вакуолярных структур. Отличительной чертой периферических отделов альвеолоцитов 1-го типа была интенсивное везикулообразование, что также отражает высокую транспортную активность данных клеток. Базаьная мембрана была разрыхлена, а в области аэрогематического барьера полностью сливалась с базальной мембранной эндотелиальных клеток. Изредка на поверхности восстановленного эпителиального слоя располагались отторгающиеся отечные клетки и кристаллические решетки сурфактантного комплекса (рис. 2).
Альвеолоциты 2-го типа также содержали крупное ядра с неровными волнистыми контурами и глубокими инвагинациями ядерной оболочки. Перинуклеарное пространство была слегка расширено. Отмечена выраженная внутриклеточная регенерация осмиофильных пластинчатых тел, которые имели различные размеры, вплоть до гигантских полостей, заполненных осмиофильным пластинчатым материалом (см. рис. 1).
Митохондрии характеризовались крупными размерами, четкими контурами митохондриальных мембран, зернистой дезорганизацией отдельных крист на фоне просветленного матрикса. Наружная митохондриальная мембрана тесно контактировала с мембранами гранулярного эндоплазматичесеого ретикулума, обильно снабженными мембрано связанными рибосомами. Гиперплазированные канальцы гранулярного эндоплазматического ретикулума были слегка расширены и заполнены хлопьевидным материалом повышенной электронной плотности.
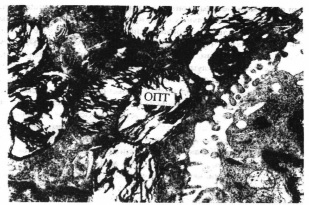
Рис. 1. Резкое увеличение количества и обьема осмиофильных пластинчатых телец (ОПТ) в цитоплазме альвеолоцита 2-го типа. Электронограмма. Ув. 21 ООО

Рис. 2. Кристаллические решетки сурфактантного комплекса (стрелка) Эпектронограмма; Ув. 18 ООО
Аппарат Гольджи также находился в состоянии гиперплазии и был представлен несколькими диктиосомами, состоящими из нескольких рядов уплощенных цистерн и многочисленных вакуолей. Апикальная поверхность альвеолоцитов 2-го типа была покрыта многочисленными микроворсинками. Наряду с высоко дифференцированными нами были отмечены альвеолоциты 2-го типа и с начальными признаками дифференцировки. Характерной особенностью данных клеток было наличие мелких гомогенных осмиофильных мультивезикулярных телец с несколькими пузырьками, вкрапленными в электронно-плотный гомогенный материал. Вдоль апикальной и базальной поверхности располагались многочисленные мелкие пиноцитозные пузырьки.
В отличие от контроля в условиях гипоксии нами отмечено появление альвеолоцитов 3-го типа, так называемых щеточных эпителиоцитов, которые имели пирамидальную форму и характерные цилиндрические микроворсинки на апикальной поверхности. Крупные митохондрии возле ядра отличались большим числом поперечно расположенных удлиненных крист. Гиперплазированные канальцы гранулярного эндоплазматического ретикулума были расширены и заполнены хлопьевидным материалом повышенной электронной плотности. Особый мембранный комплекс, множество микрофибрилл, микротрубочек и мелких осмиофильных лизосомоподобных образований свидетельствовали об интенсивности синтетических и секреторных процессов, о повышенной функциональной активности данных клеток.
Отмечено, что эндотелиоциты кровеносных капилляров, расположенные в интерстициальном слоев области слияния базальных мембран эпителия и эндотелия, превращались в истонченные пластины, лишенные органелл и рибосом, тогда как другая их поверхность их, находящаяся ближе к интерстицию, резко утолщалась, заполняясь многочисленными рибосомами, органеллами и пиноцитозными пузырьками. Среды многочисленных фибробластов различимы популяции миофибробластов и фиброкластов. Первые, вероятно, связаны со стабилизацией поверхности альвеол, вторые – с резорбцией коллагеновых фибрилл, пучки которых заполняли расширенный интерстициальный слой.
Алвеолярные макрофаги характеризовались высокой фагоцитарной и метаболической активностью. В их цитоплазме были отмечены многочисленные первичные и вторичные изосомы, фагосомы с включенным фагоцитированным пластинчатым материалом.
Таким образом, полученные нами данные свидетельствуют развитии на 15-е сутки вдыхания крысами гипоксической смеси адаптационных ультраструктурных перестроек как в эпителиальном, так и в эндотелиальном компонентах легочных альвеол в форме усиления метаболической функции. Повышалась также фагоцитарная активность легочных микрофагов. Существенную роль поддержании постоянства формы и размеров альвеол наряду с усиленным синтезом альвеолярного сурфактанта играли и гладкомышечные элементы, способствующие контракции эпителиальной выстилки альвеол.
Таким образом, полученные нами электронно-микрокопические данные подтверждают развитие выраженных адаптивных реакций в легких экспериментальных животных при многократной гипоксической гипоксии [4]. Адаптивные механизмы ултраструктурной перестройки в респираторном отделе легких затрагивали все звенья эпителиального и эндотелиального компонентов легочных альвеол в форме усиления метаболической функции. Активизировались деятельность сурфактантной системы легких, что проявлялось в резко выраженной гиперплазии и увеличении обьема осмиофильныхпластинчатов телец. Появление мелких осмиофильных пластинчатых телец в цитоплазме альвеолоцитов 1-го типа и крупных фагоцитированных пластинчатых телец в цитоплазме альвеолярных макрофагов свидетельствовало о поглощении избыточного количества незрелого сурфактанта, обильно продуцируемого альвеолоцитами 2-го типа [6]. Своеобразное строение стенки интерстициальных кровеносных капилляров отражает, с одной стороны, оптимизированный переход кислорода через тончайшие эндотелиальные пластинки, а с другой-активацию транспорта структурных и энергетических компонентов из кровеносного русла через утолщенные эндотелиоциты путем интенсивного везикулообразования к альвеолярному эпителию, где реализируются процессы усиленной внутриклеточной регенерации [7]. Существенную роль в поддержании формы и размеров альвеол играют гладкомышечные элементы интерстиция, препятствующие спадению эпителиальной выстилки. Активация альвеолоцитов 3-го типа, рассматриваемых в настоящее время как нейроэпителиальные клетки, оказывающие регулирующее влияние на функциональное состояние альвеолярного эпителия [8], вполне вероятно, связана с гипоксической нагрузкой, реализующейся через стресс-лимитирующие и интегративные системы живого организма.
Библиографическая ссылка
Сапаров К.А., Нурмухан Г.С., Султанова А.Ж. МОРФОЛОГИЧЕСКИЕ АСПЕКТЫ РЕСПИРАТОРНОГО ОТДЕЛА ЛЕГКИХ ПРИ ДОЛГОВРЕМЕННОЙ ГИПОКСИЧЕСКОЙ ГИПОКСИИ // Международный журнал прикладных и фундаментальных исследований. 2015. № 12-4. С. 677-680;URL: https://applied-research.ru/en/article/view?id=8002 (дата обращения: 01.07.2026).