

Охотничье поведение большинства наземных хищников включает ряд поведенческих реакций, связанных с ориентацией хищника в направлении добычи, сближением, контактом (нападением), обработкой, последующим умерщвлением и поеданием добычи [1]. Эти компоненты организованы в устойчивую последовательность действий – поведенческий стереотип [10]. Стереотипы охоты, как и любые другие стереотипы в поведении животных, основаны на процессах созревания врожденных программ поведения в сочетании с приобретением индивидуального и социального опыта [9].
Возможность охоты грызунов на насекомых хорошо описана в литературе [4, 6, 7]. В частности, в экспериментах по изучению взаимодействия муравьев и грызунов выращенные в лаборатории и не имевшие опыта охоты полевые мыши Apodemus agrarius успешно охотились, убивая и поедая до 36 муравьев за 10 мин [5]. Исследователи высказали предположение о том, что полевые мыши обладают врожденным стереотипом охоты на насекомых, включая и опасные их виды.
Один из универсальных вопросов при исследовании видовых стереотипов поведения заключается в выяснении вклада индивидуального опыта и созревания. Возможны различные варианты: от поведения, почти полностью основанного на индивидуальном опыте, но часто единообразного, поскольку оно сформировано сходными требованиями среды, до полностью врожденного поведения, которое проявляется либо по принципу «все и сразу», либо требует созревания [9]. Ранее для ответа на этот вопрос применялись «Каспар-Гаузер эксперименты», то есть, выращивание животных в изоляции [9]. Мы предложили метод анализа поведенческих последовательностей как «текстов», состоящих из букв – элементов поведения. Метод основан на различиях в степени сжатия текстовых файлов архиваторами и позволяет разделить врожденное поведение и приобретенные навыки без экспериментальной работы, только на основе сравнения этограмм молодых и взрослых животных. Это было показано на примере охотничьего поведения муравьев и территориального поведения чаек [8]. В данной работе мы впервые применили этот метод к грызунам, используя в качестве модели серую крысу. Этот вид известен своим гибким и разнообразным поведением как хищник-полифаг [6], и в то же время в нейрофизиологических исследованиях показано, что охотничье поведение взрослых крыс является стереотипной последовательностью действий [2]. Целью нашей работы было выявление ранее неизвестного вклада врожденного поведения и индивидуального опыта в формирование охотничьего стереотипа у серой крысы с помощью математического анализа этограмм.
Материалы и методы исследования
Исследования проводились в 2012–2014 гг., в лаборатории на беспородных серых крысах (Rattus norvegicus), не имевших опыта охоты. Половозрелые животные содержались в клетках по 3-4 особи, крысята находились в клетках вместе с матерями. Животные имели постоянный доступ к воде и пище. Кормление проводилось ежедневно после проведения тестов. В виварии соблюдался 16:8 (день-ночь) световой режим.
В исследовании приняли участие 81 половозрелая крыса (40 самок и 41 самец) в возрасте от трех месяцев до года и 52 крысенка (23 самки и 29 самцов) в возрасте от 30 до 40 дней. Тестирование начинали с этого возраста, так как предварительно на отдельной группе из семи 25-дневнх крысят было показано полное отсутствие интереса к добыче. Не охотившиеся или охотившиеся неуспешно в первом тесте крысята через 5-8 дней тестировались повторно.
Для наблюдения за процессом охоты животных по одному помещали в прозрачную арену фирмы Noldus 45×45×50 см для взрослых крыс и 30×30×35 см для крысят. Спустя 5 мин предъявляли добычу (бросали в арену сверху) и начинали фиксировать поведение с помощью видеокамеры. Тестируемым крысам предлагалось последовательно поохотиться на 3 единицы подвижной добычи – имаго мраморного таракана (Nauphoeta cinerea), средняя длина тела 27,93 ± 0,22 мм (n = 30). Наблюдения длились до завершения поедания добычи, либо, если животное не проявляло интерес к добыче, прекращались через 10 мин. Анализировались только случаи успешной охоты, закончившиеся поимкой и поеданием добычи.
Для описания поведения в качестве элементарной единицы нами выделяются элементарные двигательные акты и позы («элементы поведения»). Поведенческой последовательностью мы называем произвольный набор последовательно совершаемых элементов поведения. Внутри поведенческих последовательностей мы выделяем поведенческие стереотипы, состоящие из устойчиво повторяющихся «цепочек» элементов поведения (подробно см.: [10]). Всего было выделено 16 элементов поведения, которые разделили на 3 типа. Ключевые элементы, без которых совершение стереотипа невозможно: преследование добычи бегом (Q) или спокойным шагом (S), укус (W) и захват добычи лапами (E). Дополнительные элементы («приготовления» к охоте и поеданию добычи) присутствовали не во всех стереотипах: принюхивание (D), перенос добычи в зубах (G), перехват (R) и откусывание конечностей добычи (H). «Шумовые элементы» (не влияющие на совершение стереотипа): замирание (C), поворот корпуса на 90 ° (V), разворот корпуса 180 ° (B), поворот головы (F), стойка с опорой на арену (Y), движения назад (U) и чистка (X).
Обработка видеозаписи проводилась с 25 кратным замедлением в программе The Observer XT 10 (Noldus Information Technology). Используя полученный «алфавит» из 16 элементов, мы преобразовали демонстрируемое поведение в последовательности букв, где каждая буква соответствовала одному элементу поведения. Полученные поведенческие последовательности охотничьих стереотипов перемещались в отдельные для каждой возрастной группы «суммарные» текстовые файлы (в формате .txt).
Для парных сравнений долей успешно и неуспешно охотившихся особей и долей элементов поведения использован точный тест Фишера. Среднее количество различных элементов поведения в стереотипах сравнивали t-критерием Стьюдента. Для визуализации структуры стереотипа, на основании «суммарных» текстовых файлов, рассчитывались матрицы вероятностей перехода от одного поведенческого элемента к другому (Марковский процесс первого порядка), которые использовались для построения схемы стереотипа.
Оценка связанности между элементами поведения в поведенческих последовательностях охотничьих стереотипов проводилась с помощью метода анализа биологических текстов на основе идей Колмогоровской сложности и проверки статистических гипотез [8]. Для этого нами написана программа, которая случайным образом выбирала из «суммарных» файлов целые последовательности и переносила их в отдельный текстовый файл заданного размера. Получено по 5 файлов объемом 300, 400 и 500 байт содержащих стереотипы взрослых крыс и крысят. При этом мы следили за тем, чтобы каждая последовательность не была скопирована дважды (то есть присутствовала только в одном файле). Полученные файлы сжимались при помощи архиватора 7-zip с использованием метода сжатия BZip2. Проводилась сравнительная оценка степеней сжатия успешных охотничьих стереотипов (подробно см.: [10]). Под степенью сжатия понимается отношение размера заархивированного файла к его исходному размеру. Чем меньше степень сжатия, тем меньше сложность записанных в текстовом файле поведенческих стереотипов, и наоборот, чем степень сжатия выше, тем они сложнее.
Результаты исследования и их обсуждение
При первом предъявлении подвижной добычи охотничье поведение проявили 14 крысят в возрасте 30 дней, 6 животных в возрасте 32 дней, 2 крысенка в возрасте 33 дня, 8 особей в 34 дневном возрасте и 55 взрослых крыс. Доли молодых 57,7 % (30 из 52) и взрослых животных 67,9 % (55 из 81), охотившихся в первом тесте, достоверно не различалось (точный тест Фишера, p = 0,2689). После повторного тестирования не охотившихся или охотившихся неуспешно крысят суммарная доля успешных молодых охотников составила 88,5 % (46 из 52), что достоверно больше, чем у взрослых 67,9 % (55 из 81) (p = 0,007). Молодые животные продемонстрировали меньшее количество неуспешных охот (40 из 159), чем взрослые (77 из 202) (p = 0,009). Стоит отметить наличие успешных охотников, которые ни разу из трех охот не теряли добычу: 23 взрослых крысы и 20 крысят.
Длина успешных охотничьих стереотипов 30-дневных крысят составила 45,4 ± 6,5, у 34-дневных 33,8 ± 4, у 40-дневных крысят 32,1 ± 4 элемента поведения (достоверных различий не выявлено). Схемы успешных охотничьих стереотипов у этих трех возрастных групп оказались сходны (рис. 1, а). Поэтому мы объединили все полученные стереотипы молодых животных в одну группу. В целом успешные охотничьи стереотипы крысят оказались длиннее 35,2 ± 2,3, чем у взрослых животных 21,9 ± 1,7 (t = 4,7, p < 0,01).
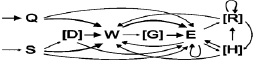
а)
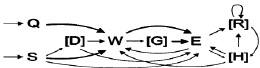
б)
Рис. 1. Схемы охотничьих стереотипов крысят (а) и взрослых крыс (б). Тонкой пунктирной линией обозначены некоторые неустойчивые связи между элементами (p < 0,2). Простой линией обозначены устойчивые связи (0,2 ≤ p < 0,5). Жирной линией обозначены высоко устойчивые связи между элементами (p ≤ 0,5). В квадратных скобках указаны дополнительные элементы
На основании рассчитанных матриц вероятностей переходов между элементами поведения в поведенческих последовательностях построены схемы охотничьего стереотипа в отношении подвижной добычи (рис. 1). Следует отметить, что на схеме показаны все устойчивые связи между элементами (p < 0,2) и некоторые неустойчивые (p < 0,2), но важные для совершения стереотипа связи.
У молодых и взрослых животных стереотип, как правило, начинался с преследования добычи бегом (Q) или спокойным шагом (S), а в тех редких случаях, когда добыча сама приближалась слишком близко, охотничий стереотип мог начинаться без фазы преследования, затем могло следовать принюхивание (D). Далее, как правило, следовал укус (W) и захват добычи в передние лапы (E). Реже крысы и крысята могли начать атаку с одной или нескольких попыток захватов добычи передними лапами. Такие повторяющиеся цепочки элементов поведения «захват лапами – захват лапами» присутствовали в 83,2 % (99 из 119) успешных охотничьих стереотипах крысят, но лишь в 49,6 % (62 из 125) стереотипах взрослых (p = 2,352*10-8). После захвата добычи передними лапами, чаще всего наблюдались перехваты удерживаемой добычи (R). В дальнейшем мог наблюдаться акт откусывания конечностей добычи (H). Последний элемент поведения, предшествующий поеданию, выступает в качестве окончания успешного охотничьего стереотипа, ими могли быть: укус (W), захват лапами (E), перехват (R) или откусывание конечностей добычи (H).
В успешных охотничьих стереотипах крысят доли шумовых (p = 0,0269) и дополнительных элементов (p = 0,0096*10-3) достоверно ниже, а ключевых элементов достоверно выше (p = 0,0025*10-8), чем у взрослых крыс. Количественный анализ проводился для ключевых элементов поведения и наиболее часто встречающихся дополнительных: «перехват», «откусывание конечностей добычи». Результаты представлены на рисунке 2. Крысята чаще чем крысы преследовали свою добычу «бегом» (t = 5,6, p < 0,01). На один успешный охотничий стереотип у крысят приходилось больше элементов поведения «укус» и «захват лапами», чем у взрослых крыс (t = 5,3, p < 0,01; t = 5,5, p < 0,01 соответственно). В одном охотничьем стереотипе крысенка в среднем встречалось больше связок элементов поведения «укус – захват лапами», чем у взрослых крыс (t = 4,3, p < 0,01). Среднее количество дополнительных элементов поведения «преследование шагом» (t = 0,75, NS), «перехват» (t = 1,5, NS) и «откусывание конечностей добычи» (t = 1,1, NS) в стереотипах крыс и крысят не различалось.
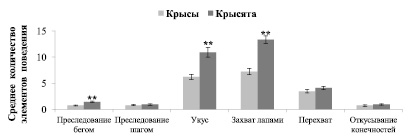
Рис. 2. Среднее количество элементов поведения в одном стереотипе
Степени сжатия файлов содержащих успешные охотничьи стереотипы взрослых крыс и крысят достоверно не различались при объемах файлов 300, 400 и 500 байт (Критерий Манна-Уитни, Uэмп = 12; Uэмп = 8; Uэмп = 5; Uкр = 1, NS) (рис. 3).
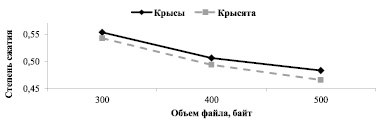
Рис. 3. Различия в степенях сжатия файлов разного объема
Проявление охотничьего стереотипа у взрослых и молодых крыс носит факультативный характер, то есть, встречается не у всех особей. У крысят стереотип проявляется в возрасте 30 – 34 дней. Индивидуальная вариабельность и возраст проявления охотничьего стереотипа у крыс оказались сходными с полифагом сирийским хомячком (Mesocricetus auratus) [7]. Это отличает полифагов от специализированных хищных грызунов – северного и южного кузнечикового хомячка (род Onychomys), у которых охотничий стереотип в отношении сверчков проявляется уже в 22-дневном возрасте, а его проявления у «наивных» животных носят облигатный характер [4].
Большинство «наивных» взрослых крыс охотились успешно, что свидетельствует об отсутствии критического периода созревания стереотипа. Атака на добычу начиналась c преследования, укуса, за которым следовал захват добычи лапами. Аналогичным образом (укус – захват лапами) атакуют добычу (сверчков) сирийские хомячки [7]. Хищные кузнечиковые хомячки, наоборот, чаще начинают свою атаку на сверчков с захвата добычи передними лапами, после чего начинают ее кусать [5]. Считается, что начало атаки с захвата добычи в передние лапы является прогрессивной эволюционной чертой, а начало атаки с укуса – более древняя и примитивная форма [3]. Именно она оказалась характерной для серой крысы. В схемах стереотипов крысят и крыс выявлено несколько различий. Наличие связи между элементами поведения «преследование бегом» и «захват лапами» у крысят обусловлено тем, что они значительно чаще, чем взрослые, преследовали свою добычу бегом. Повторяющиеся серии захватов добычи лапами в стереотипах крысят мы связываем с «неуклюжестью» крысят, которым приходится повторять попытку захвата. Связь между элементами «перехват добычи» и «укус» также можно объяснить «неловкостью», заключающейся в манипуляциях с сопротивляющейся добычей, удерживаемой передними лапами. В результате добыча падает на дно арены и животному вновь приходится ее преследовать или атаковать (кусать, захватывать лапами). При этом общий порядок совершения поведенческих актов остается неизменным: преследование, укус, захват лапами и последующие манипуляции (перехваты) и обработка добычи (откусывание конечностей). Отсутствие устойчивой связи между элементами поведения «захват лапами» и «перехват добычи» связанно с тем, что вероятность переходов между повторяющимися «захватами лапами» выше, и снижается вероятность перехода к другим элементам. В стереотипах крыс и крысят содержалось близкое количество «перехватов», что в совокупности с необходимостью предварительного «захвата добычи лапами» подтверждает связанность этих элементов поведения. Интересно также отметить, что в стереотипах молодых и взрослых животных элементы поведения «перехват добычи» были организованны в повторяющиеся цепочки (перехват – перехват). Несмотря на незначительные различия в схемах стереотипа крыс и крысят, порядок совершения ключевых элементов поведения в них одинаков.
Результаты сравнительного анализа сложности успешных охотничьих стереотипов, основанные на применении понятия Колмогоровской сложности [9] показали, что средняя степень сжатия стереотипов взрослых крыс и крысят значимо не различалась. Для лучшего понимания этого результата следует пояснить основные принципы, лежащие в основе использованного метода сравнения сложности. Программа – архиватор находит в последовательностях букв, записанных в текстовом файле, закономерности (повторяющиеся участки), и чем больше и длиннее будут такие участки, тем в результате лучше будет сжиматься такой файл (степень сжатия будет меньше). С другой стороны, в стереотип могут вклиниваться случайные («шумовые») элементы поведения. Они не виляют на совершение стереотипа, но понижают упорядоченность последовательности букв, и тем самым увеличивают степень сжатия. Сложность стереотипов крыс и крысят не различалась, значит, уровень связанности между элементами поведения у них близок, и устойчивость связей, отраженных в схемах стереотипов, одинакова.
Различия в длинах охотничьих стереотипов взрослых и молодых крыс обусловлены большей долей ключевых элементов поведения в стереотипах крысят. Это подтверждается существенными различиями в среднем количестве ключевых элементов «укус», «захват лапами» у взрослых и молодых особей, тогда как количество дополнительных элементов «перехват» и «откусывание конечностей» у них не различается. Отметим, что тесные связки элементов «укус – захват лапами», чаще встречались в стереотипах крысят. Мы предполагаем, что многочисленные повторы связок этих элементов обусловлены процессом оптимизации навыка захвата добычи.
Таким образом, анализ этограмм охотничьего поведения с помощью метода, основанного на идеях Колмогоровской сложности, показал, что у молодых и взрослых крыс порядок совершения элементов поведения и уровень их связанности сходен. У «наивных» животных стереотип охоты проявляется по принципу «все и сразу». В совокупности эти данные позволяют предполагать врожденный характер охотничьего стереотипа у крыс. Критический период созревания стереотипа отсутствует. Оптимизация охоты у молодых особей достигается за счет совершенствования отдельных элементов поведения и связок между ними.
Исследования поддержаны Российским научным фондом (грант № 14-14-00603).
Библиографическая ссылка
Левенец Я.В., Пантелеева С.Н. СРАВНИТЕЛЬНЫЙ АНАЛИЗ ОХОТНИЧЬЕГО ПОВЕДЕНИЯ У ВЗРОСЛЫХ И МОЛОДЫХ СЕРЫХ КРЫС // Международный журнал прикладных и фундаментальных исследований. 2016. № 8-5. С. 741-746;URL: https://applied-research.ru/ru/article/view?id=10160 (дата обращения: 01.07.2026).