

Европейский хариус (Thymallus thymallus), как и все лососевые рыбы, обладает ценными вкусовыми качествами, поэтому в последнее время растет интерес к хариусу не только как объекту любительского лова, но и виду, важному для аквакультурного разведения. В некоторых регионах России численность природных популяций европейского хариуса сокращается и этот вид, являясь не только объектом хозяйственного интереса, но и важным звеном водных экосистем, внесен в Красную книгу этих регионов. Рациональное использование, искусственное разведение и охрана вида невозможны без применения современных генетических методов. При этом хариус является объектом не только прикладных, но и фундаментальных исследований. Одним из таких важных направлений являются филогенетические исследования, на базе которых мы можем уточнить положение вида в современной систематике и предположить пути его расселения. Несмотря на то, что в последнее время в научной литературе интерес к этому виду достаточно высок [3–5, 7–9], в российской части ареала большинство популяций остаются неисследованными, данные фрагментарны и затрагивают отдельные реки или их притоки [3, 5, 7, 9].
Соответственно, в задачи нашего исследования входило: уточнить филогенетическое положение европейского хариуса на территории Европейского Севера, а также предположить возможные пути его расселения. Для выполнения этой задачи в работе анализируется контрольный регион мтДНК, относящийся к некодирующим последовательностям мтДНК и относительно быстро накапливающий замены.
Материалы и методы исследования
Всего было проанализировано 60 особей хариуса из разных рек бассейна Белого моря. Географическое расположение выборок показано на рис. 1. Выделение ДНК из плавников проводили с использованием набора QIAGEN DNeasyTM по протоколу производителя. Для амплификации фрагментов использовали последовательности праймеров LRBT–25, LRBT–1195 [8]:. Амплификацию проводили в 15 мкл смеси, содержащей 10ХTaq буфер, 2 мМ Mg2+, 2.5 мM dNTP’s, 1ед. Taq полимеразы (ДИАЛАТ, Россия), 5 пкM каждого праймера, 1.5 мкл ДНК. Амплификацию проводили в термоциклере Tetrad 2 (Bio–Rad Laboratories, Inc., США) в следующем режиме: I – 95 °С 5 мин., затем 35 циклов II – 94 °С 20 сек., 30 сек. 58 °С, 72 °С 1мин., заключительная элонгация 72 °С – 10 мин. Анализ нуклеотидной последовательности проводился в генетическом анализаторе ABI PRISM 3500 (Applied Biosystems, США). Первичная обработка и множественные выравнивание нуклеотидных последовательностей проводили в программе Geneious® 6.0.5, статистический и филогенетический анализы проводили в программах DnaSP V.5 [6], Geneious® 6.0.5 (Biomatters Ltd.). Для построения дендрограммы были использованы последовательности контрольного региона мтДНК, депонированные в базе нуклеотидных последовательностей GenBank® (NCBI).
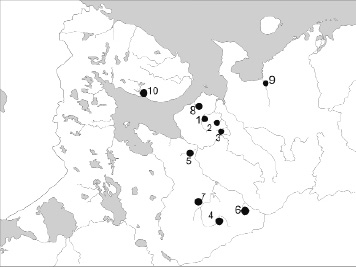
Рис. 1. Карта расположения выборок. Притоки р. Кулой: 1 – Сояна, 2 – Лака, 3 – Келда; притоки р. Северная Двина: 4 – Устья, 5 – Обокша, 6 – Кестваж, 7 – Леменьга; 8 – р. Мегра; 9 – р. Ома; 10 – р. Варзуга
Результаты исследования и их обсуждение
После множественного выравнивания полученных последовательностей исследован участок мтДНК длиной 1046 п.н., включающий полную последовательность контрольного региона Д–петли, GC–содержание этого фрагмента составляло 35.6 %. Было выявлено 13 полиморфных сайтов (1,2 % от всех сайтов), из них информативных 9. Из 14 замен 1 трансверсия, остальные транзиции, также выявлена одна делеция. Среди исследованных последовательностей выделено 10 гаплотипов, 8 – впервые. Распределение гаплотипов по выборкам представлено в таблице.
Распределение гаплотипов в исследованных выборках хариуса
|
Выборки |
Hap1 Main |
Hap2 Ku1 |
Hap3 Ku2 |
Hap4 Ku3 |
Hap5 Meg |
Hap6 ND1 |
Hap7 Ku4 |
Hap8 Ku5 |
Hap9 ND2 |
Hap10 Var |
|
река Кулой |
||||||||||
|
р. Келда |
8 |
2 |
||||||||
|
р. Лака |
14 |
1 |
1 |
1 |
||||||
|
р. Сояна |
11 |
1 |
||||||||
|
река Северная Двина |
||||||||||
|
р. Устья |
1 |
|||||||||
|
р. Обокша |
2 |
|||||||||
|
р. Кестваж |
2 |
6 |
||||||||
|
р. Леменьга |
1 |
1 |
||||||||
|
река Мегра |
2 |
1 |
||||||||
|
река Ома |
1 |
|||||||||
|
река Варзуга |
4 |
Гаплотип, обозначенный нами «Main», выявлен в большинстве выборок за исключением выборки из реки Варзуга. В реке Кулой было обнаружено 6 гаплотипов, 3 из которых являются вариантами основного гаплотипа с однонуклеотидными заменами (рис. 2), а 2 других гаплотипа (Ku4 и 5) с большим количеством замен. Гаплотип Ku5 соответствует гаплотипу AF522430 (At6) (рис. 3), обнаруженному в выборке из бассейна Балтийского моря [9]. В реке Северная Двина было выявлено 3 гаплотипа, гаплотип ND1 относится к группе массового гаплотипа «Main»с одной заменой, а гаплотип ND2 на 5 замен отличается от массового гаплотипа «Main». Гаплотип ND2 (рис. 3) аналогичен гаплотипу AF522426 (At2) выявленному в реке Тана на севере Норвегии [9]. В популяции реки Мегра присутствует гаплотип Meg, отличающийся от основного на 2 замены. В реке Варзуга обнаружен только 1 гаплотип Var, характерный только для рыб из этой реки, отличающийся от массового гаплотипа на 7 замен (рис. 2) и относящийся к той же группе гаплотипов, что и ND2 (рис. 3).
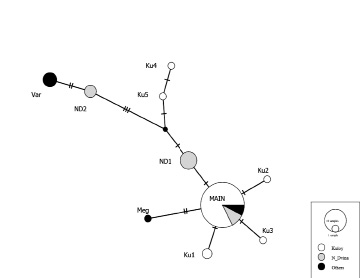
Рис. 2. Сеть гаплотипов контрольного региона мтДНК исследованных рыб на основе алгоритма TCS [2]. Белым цветом выделен Кулой, серым – Северная Двина, черным – выборки из других рек. Обозначения гаплотипов: Ku – Кулой, ND – Северная Двина, MAIN – наиболее распространённый гаплотип, Meg – Мегра, Var – Варзуга
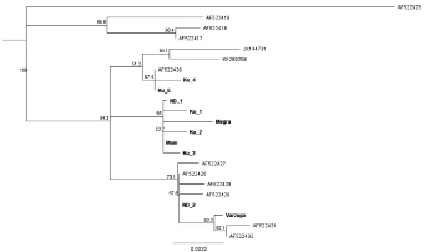
Рис. 3. Филогенетическое дерево гаплотипов контрольного региона мтДНК североатлантической группы европейского хариуса. Дерево построено на основе алгоритма программы Mr Bayes 3.2.1. методом присоединения соседей с использованием HKY модели, в качестве корня использован гаплотип из реки Луары (AF522425). В узлах дерева указаны значения бутстрепа. На дендрограмме гаплотипы, полученные в нашей работе, обозначены Ku_ – Кулой, ND_ – Северная Двина, Main – наиболее массовый гаплотип
На основе проведенного анализа полученные нами гаплотипы относятся к выделенной в предыдущих исследованиях скандинавской филогенетической линии европейского хариуса и образуют три клады, предположительно отражая рефугиумы, из которых происходило расселение хариуса (рис. 3). Во время максимального оледенения северные реки, по мнению Гросвальда [1], сбрасывали воды на юг, соответственно, формирование русел рек Европейского Севера происходило после отступления ледника и расселение хариуса происходило по мере формирования русел. Наиболее вероятно, в период покровного оледенения вид мог сохраняться в сети приледниковых озер, служивших рефугиумами для вида. Большинством исследователей [4, 7] принято допущение, что у холодноводных видов рыб различия в Д–петле достигают 1 % за 1 млн лет. Различия между гаплотипами из разных клад в нашем исследовании достигают от 0.4 % до 0.9 %, при этом клада с гаплотипами из Варзуги (Var) и Северной Двины (ND2) c высокой бутстреп поддержкой (~ 70 %), что предполагает разделение вида и существование вида во время последнего максимального оледенения по крайней мере в двух рефугиумах. Мы можем предположить существование западного рефугиума, откуда происходило заселение хариусом водоемов Фенноскандии и северо-западных притоков Северной Двины. Другим возможным местом рефугиума мог быть расположенный на юго-востоке, возможно, в районе существовавшего в максимальной стадии последнего оледенения Печорского приледникового озера, откуда пошло расселение хариуса в восточные и северо-восточные реки. В пользу этой гипотезы свидетельствует то, что массовый гаплотип обнаружен во всех исследованных популяциях северо-восточных рек и притоков Северной Двины, тогда как в Варзуге и других реках Фенноскандии этот гаплотип и его варианты обнаружены не были. Судя по полученным результатам, предполагаемый рефугиум для популяций Камы и Урала также отличался от этих двух, так как там тоже не был обнаружен гаплотип, наиболее часто встречающийся в наших выборках, а также эти гаплотипы, выявленные в работе Марича с коллегами [7], достаточно сильно (~ 0.5 %) отличаются от остальных гаплотипов скандинавской линии. Для более точного понимания картины расселения европейского хариуса на территории Европейского Севера необходимо более полное исследование выборок не только из рек Арктического побережья, но и притоков верхней Волги.
Заключение
В работе проанализирован фрагмент мтДНК, включающий полную последовательность контрольного региона, у 60 особей европейского хариуса из выборок рек бассейна Белого моря. Было выявлено 10 гаплотипов, 8 из которых были описаны впервые. На основе проведенного анализа все гаплотипы относятся к скандинавской филогенетической линии и образуют три клады, одна из которых с высокой бутстреп поддержкой, что позволяет предположить расселение европейского хариуса на территории Европейского Севера по крайней мере из двух рефугиумов.
Библиографическая ссылка
Пономарева Е.В., Пономарева М.В., Шубина Е.А. РАЗНООБРАЗИЕ ГАПЛОТИПОВ КОНТРОЛЬНОГО РЕГИОНА МТДНК ЕВРОПЕЙСКОГО ХАРИУСА (THYMALLUS THYMALLUS L.) РЕК БАССЕЙНА БЕЛОГО МОРЯ // Международный журнал прикладных и фундаментальных исследований. 2016. № 8-5. С. 747-751;URL: https://applied-research.ru/ru/article/view?id=10161 (дата обращения: 01.07.2026).