

Наследуемость продуктивности у сосны изучалась нами в рамках проекта по выведению сортов хвойных пород для плантационного выращивания, начатого в 1979 г. Написан ряд крупных работ [5-8]. В результате удалось выяснить, что эффект улучшения семян на лесосеменных плантациях (ЛСП) и на постоянных лесосеменных участках (ПЛСУ) возникает вследствие более комфортных условий формирования семян и выражается в усилении роста культур в первые 12 лет на 5–6 %. Непременным условием такого эффекта является точное соответствие эдатопов; например, если ЛСП заложены в условиях свежей субори В2, то и культуры должны выращиваться в ней же. Отклонение даже на одну градацию эдатопа резко снижает эффект. При этом собственно «генетическое» улучшение таких семян остается неизвестным [5, 6].
Лесная селекция находится в начале своего развития и до сих пор точно неизвестно, какова доля влияния генотипа, среды и взаимодействия генотип-среда у разных пород. Приблизительно считают [6], что у сосны обыкновенной сила влияния генотипа на рост деревьев не превышает 5–10 %. Остальное – это влияние среды (≈ 60 %) и взаимодействие генотип-среда (≈ 35 %), которые сейчас относят к эпигенетике [3]. Как же с ней обстоят дела? В лесной селекции она открывает массу новых возможностей, однако исследования по ней единичны. Появились сведения и о неизвестных ранее явлениях, например, геоактивных зонах Земли [7] и фитогенных полях деревьев [2], изучение которых позволяет находить и учитывать целый ряд новых факторов, влияющих на наследуемость.
Цель работы – в родительских и дочерних насаждениях изучить влияние почвенных условий и геоактивных зон Земли на наследуемость роста у сосны обыкновенной.
Материалы и методы исследования
Объектом исследования были 123 дерева сосны обыкновенной (Pinus sylvestris L.) в возрасте 26 и 55 лет, растущие ПЛСУ. Участок сформирован изреживанием культур, созданных на старой пашне в 1962 г. посадкой сеянцев по схеме 2.0×1.0 м. Почва супесчаная, подстилаемая с 90–120 см плотным суглинком. Местоположение: юго-восточный склон 3 ° пологого холма. Тип условий сосняк кисличник (эдатоп B2). Культуры в возрасте 10 лет изредили: вырубили 2 ряда из трех с оставлением лучших растений в рядах через 5–8 м. Затем в возрасте 18 лет у 60 % деревьев обрезали кроны, удаляя 5–6 приростов. В 22 года проведена оценка семеношения и собран урожай от 234 деревьев, затем у них измеряли высоты. Спустя 29 лет в возрасте 55 лет у деревьев измеряли диаметр ствола. Координаты участкаN: 58˚3΄57΄΄, E: 56˚ 2΄36΄΄, кв. 18 Нижне-Курьинского лесничества, леса г. Перми.
Потомство первого урожая (1982 г.) выращивали в грунте и в теплице по технологии многообъемных испытаний [6]. Сеянцы сортировали и отбирали 50–70 % лучших, из которых 60 шт. высаживали в школу на 2 года. Эдатоп школы совпадал с эдатопом ПЛСУ. Затем у 3-летних саженцев измеряли высоты и планировали их посадку в тест-культуры. Из-за дефицита площади заложили только один участок площадью 1.5 га в кв. 83 (ныне 65) Нижне-Курьинского лесничества с высадкой 123 семей из 234. Для этого отобрали семьи с высотами от 97-100 %, сам контроль и часть слабо растущих семей. Эдатоп здесь также совпадал с эдатопом ПЛСУ. Измерения высот проведены в 9, 12 и 18 лет.
Потомство второго урожая (1985 г.) выращивали в питомнике в эдатопе В2 до двух лет и далее высадили в культуры в эдатопе А2 на вырубке площадью 7 га в кв. 29 Оханского лесничества. Здесь была другая проблема – культуры сосны в Пермском крае в те годы повреждали лоси. Удалось сохранить эти культуры только до 9-летнего возраста.
Для оценки роста матери использован диаметр ствола, для оценки потомства – средняя высота 20–70 растений (высота семьи) в % от контроля. Контролем служили семена из 5 лесхозов. Всего было измерено от первого урожая 6140 и 2360 растений в возрасте 3 и 18 лет, от второго урожая 5388 растений в возрасте 9 лет и 987 растений в контроле.
В качестве методов использованы обычные методы таксации и статистический метод обработки данных, а также биолокационный метод [2, 4, 7], с помощью которого вблизи материнских деревьев определяли геоактивные зоны пяти видов и их диаметры, в т.ч. широко известные патогенные зоны Хартмана и Карри. Библиография по этой теме превышает три сотни работ, однако изучению растений посвящены единицы, так как всех интересует в основном их влияние на человека. Нами взята за основу методика В.А. Поносова [4] и наш опыт работы, изложенный в монографии [7]. В основу метода положена способность человека воспринимать сверхслабые поля электромагнитной, гравитационной и иной природы. В этом методе используется «прибор» под названием Человек. И если оператор провел его юстировку, то он доверяет ему и исследует окружающий мир. Биолокацию сравнивают со своеобразным талантом, но скорее это способность человека правильно (нейтрально) обращаться к своему телу, как к «биоприбору», что позволяет фиксировать неосознаваемые психикой ощущения, используя в качестве стрелки в этом «приборе» рамку, маятник или сенсор. Геоактивные зоны ныне определяют также и объективными методами контроля, фиксируя их излучения, например, на фотоматериалы, и они подтверждают данные биолокации. При этом излучения даже наиболее сильных сетей Хартмана настолько слабы, что их отображение даже на самые чувствительные фотоматериалы происходит лишь через 15 суток [1]. Поэтому их изучение будет наиболее результативно в древостоях, где деревья десятилетиями подвергаются их воздействию [2, 7].
В качестве условных обозначений использованы следующие символы: n – объем выборки, шт.; x – среднее значение; ± б – стандартное отклонение; ± m – ошибка среднего выборочного значения; t – критерий различия; r – коэффициент корреляции.
Результаты исследования и их обсуждение
В лесной селекции наследуемость роста определяют по высоте потомства на ранних этапах онтогенеза. В самом простом случае ее рассчитывают через корреляцию между размерами ствола матери и высотой семьи. Такие корреляции мы рассчитали на двух участках тест-культур в возрасте 18 и 9 лет. На первом участке условия соответствует свежей субори (B2) и они совпадают с родительскими, на втором участке условия более сухие (А2). Общая картина наследуемости на них выглядит совершенно по-разному (рис. 1).
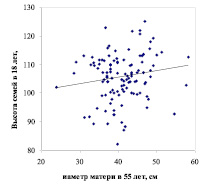
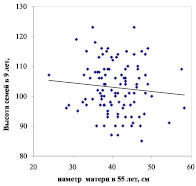
Рис. 1. Диаметры матерей и высота семей сосны при выращивании потомства в совпадающих (слева) и в несовпадающих (справа) с родительским ценозом лесорастительных условиях
На первом участке в условиях, совпадающих с материнскими, наследуемость оказалась положительной. На втором же участке, в возрасте семей 9 лет и при несовпадении условий дочернего и родительского ценозов, она стала отрицательной. Связи слабые (r = 0.152 ± 0.092 и r = – 0.097 ± 0.094), но различия между ними достоверны при t = 1.89 > t0.10 = 1.66. Из этих графиков следует, что крупные матери дают в целом более высокое потомство только при его выращивании в условиях, совпадающих с родительскими. Все это сопровождается разбросом данных и выглядит скорее как тенденция. Ее мы выявили уже достаточно давно в потомстве 1.3 тыс. деревьев сосны в 17 опытах, где испытывалось 43 тыс. потомков [6]. По сути, тенденция эта вполне в духе идей эпигенетики, т.е. в совпадающих условиях продуктивность сосны наследуется, а в несовпадающих условиях этого не происходит. Столь важные детали ускользали прежде от нашего внимания, когда мы усредняли все полученные данные о росте потомства – для суждения о генотипе матерей.
Далее мы попытались выяснить, как меняются корреляции при нахождении матерей на тех или иных геоактивных зонах. У пяти типов геоактивных зон с помощью биолокации были определены их диаметры, причем для патогенных зон Хартмана и Карри учитывали только усиленные зоны с диаметрами 0.55, 1.0 и 2.0 м, а для трех типов благоприятных зон – зоны с диаметрами 1.0, 3.0 и 8.0 м [7]. Заметим, что в «чистом виде» группы матерей по этим зонам сформировать не удалось, так как многие деревья находилась сразу в нескольких зонах, и матери из группы с патогенными зонами (кратко минус-зонами) находились, например, также и на зонах с благоприятным влиянием (плюс-зонах). Матерей разделили на пять групп: 1 – на нейтральных местах; 2, 3, 4 – на местах с плюс-зонами разного диаметра; 5 – на местах с минус-зонами в сочетании с плюс-зонами. Для этих групп получили 14 корреляций, показывающих уровень наследуемости у сосны в разных условиях (таблица).
Наследуемость продуктивности у сосны в лесорастительных условиях В2 и А2 при формировании матерей на геоактивных зонах разного типа
|
Показатели связи |
Нахождение матери на местах |
Общая выборка |
Выборка без зон 8 м |
||||
|
нейтральных |
с плюс-зонами |
с минус-зонами |
|||||
|
1 м |
3 м |
8 м |
|||||
|
Потомство 18 лет в условиях В2 (почвенные условия совпадают с родительскими) |
|||||||
|
число пар |
13 |
50 |
63 |
47 |
20 |
122* |
75 |
|
корреляция |
0,08 |
0,11 |
0,10 |
-0,06 |
0,31 |
0,15 |
0,28 |
|
ошибка |
± 0,28 |
± 0,14 |
± 0,12 |
± 0,15 |
± 0,20 |
± 0,09 |
± 0,11 |
|
достоверность |
0,29 |
0,77 |
0,81 |
0,40 |
1,54 |
1,73 |
2,59 |
|
Потомство 9 лет в условиях А2 (почвенные условия не совпадают с родительскими) |
|||||||
|
число пар |
11 |
47 |
59 |
41 |
20 |
112* |
70 |
|
корреляция |
-0,16 |
0,10 |
0,04 |
-0,48** |
0,47** |
-0,10 |
0,13 |
|
ошибка |
± 0,29 |
± 0,14 |
± 0,13 |
± 0,12 |
± 0,18 |
± 0,09 |
± 0,12 |
|
достоверность |
-0,56 |
0,70 |
0,30 |
3,93 |
2,59 |
1,04 |
1,12 |
Примечание. * – зоны перекрывают друг друга, и число пар в выборках больше их числа в общей выборке; ** – значение корреляции достоверно.
Из данных таблицы следует, что достоверные (и прямо противоположные) корреляции наблюдаются только в двух группах матерей: на патогенных зонах (r = 0.47 ± 0.18), и на благоприятных зонах 8 м (r= – 0.48 ± 0.12). Эти корреляции являются экспериментальным фактом, и их противоположные значения как-то следует объяснить. Получается, что в каких-то местах территории происходили некие многолетние воздействия на материнские деревья со стороны геоактивных зон, которые привели в одном случае к появлению положительной наследуемости, а в другом случае наоборот, наследуемость оказалась отрицательной. Поэтому мы провели имитацию отбора матерей с учетом этого нового фактора и исключили из общей выборки матерей на зонах 8 м (последняя графа в таблице). Корреляции сразу выросли вдвое: в 18 лет от 0.15 до 0.28, а в 9 лет с – 0.10 до 0.13. Это дает надежду на существенное повышение наследуемости у сосны, если материнские деревья отбирать вне территорий с благоприятными зонами диаметром 8 м.
Далее мы обнаружили, что на снижение наследуемости на зонах 8 м в условиях А2 решающее влияние оказали 9 семей с высотами менее 96 % и их крупные матери; эти семьи находятся внизу и отделены горизонтальной линией (рис. 2).
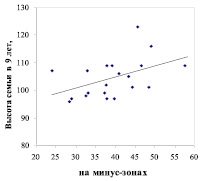
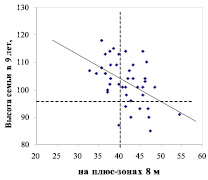
Диаметр матери, см
Рис. 2 – Диаметры деревьев сосны на патогенных (слева) и благоприятных (справа) зонах геобиологических сетей и их связь с высотой 9-летнего потомства
Выяснилась и еще одна особенность этих нежелательных для плюсовой селекции матерей, «тянущих» наследуемость в отрицательную сторону. Оказалось, что все 9 матерей со слабым потомством формировались сразу на двух типах зон с диаметрами 8.0 и 3.0 м. Т. е. зоны эти накладывались друг на друга, или они находились рядом и обе влияли на дерево. Подобное сочетание зон наблюдалось частично и у матерей, которые произвели самые быстрорастущие семьи (на правом графике верхние 7 точек), однако это случилось у них лишь 2 раза из 7 случаев. Конечно, для каких-либо категоричных выводов данных пока мало. Однако и одна выборка может дать подсказку для поиска.
По наблюдениям на множестве других объектов, именно на зонах 8 м деревья развивают иногда очень широкие, густые, а также плакучие кроны, имеют очень толстые сучья, густое охвоение и другие морфологические отклонения в максимальную сторону. Поэтому на зоны 8 м следует обратить особое внимание. Возможно, их энергия включает в работу «молчащие» гены, растения проявляют некие скрытые возможности генотипа и активизируют их, и они плохо влияют затем на рост потомства [7].
Полученные здесь результаты, с учетом ранее полученных нами данных на потомстве ели финской (Рiceafennica [Regel] Кom.) [5], позволяют предполагать, что потомство в некотором роде «помнит» и «знает», какие условия для него подходят более всего. Иными словами, в потомстве действуют некие эпигенетические механизмы; они побуждают потомства расти лучше или хуже в зависимости от того, совпадают или нет условия их развития с родительскими. Эта картина событий совершенно не вписывается в идеологему плюсовой селекции. В литературе об «отрицательной» наследуемости прямо не сообщается, но есть косвенные тому свидетельства в республике Коми [9], где в одном опыте среди потомства 129 плюс-деревьев превышали контроль в 9-летнем возрасте лишь 3 % семей, а все их потомство росло на 12.7 % хуже контроля; в других опытах в возрасте 5–18 лет превышения семей 190 плюс-деревьев колебались от нуля до минус 3.6 %.
Полученные нами результаты позволяют внести некоторую ясность в эту проблему. В частности, одной из причин снижения наследуемости может быть воздействие на материнское растение со стороны геоактивных зон Земли, энергию которых технические приборы пока не фиксируют и их определение возможно только биолокацией. Для этого метода, безусловно, нужна строгая верификация, т.е. проверка экспериментальных фактов, установленных с его помощью. Мы предложили схемы опытов в лесных культурах, реализуемых в течение 5–7 лет [7, с. 164], и пока эти опыты никем не поставлены. Они реализуемы практически на любой вырубке или на зарастающем лесом поле. Их результаты будут зафиксированы статистически через средние размеры деревьев, что позволит выяснить стимулирующее или ингибирующее влияние геоактивных и, наконец, разрешить бесполезные теоретические споры о правомерности применения биолокации.
На наш взгляд, особенно интересно резкое повышение наследуемости до достоверных значений (r = 0.47 ± 0.18) у родителей на патогенных зонах сетей Хартмана и Карри, при одновременном их нахождении также и внутри благоприятных зон. Этот факт позволяет надеяться, что если мать, испытывая одновременно и благоприятное, и повреждающее воздействие геоактивных зон, все-таки формирует при этом крупный ствол, то мы можем рассчитывать на получение от нее и хорошего потомства. Мать как бы «настраивает» свое потомство, на основе испытанных ею стрессов, на более устойчивый рост в иных (в нашем случае более сухих), чем это было у нее, почвенных условиях. Во всяком случае, именно в таком ключе эпигенетика рассматривает адаптацию материнских популяций и влияние отличительных особенностей ее генома на развитие потомства [3].
Заключение
Отрицательная наследуемость роста у сосны вызывается двумя причинами. Первая – это несовпадение эдатопов, причем всего лишь на одну градацию: B2 в материнском и А2 в дочернем насаждении. Вторая причина связана с формированием матерей на геоактивных зонах определенного типа, вследствие чего наследуемость снижается до отрицательного значения. Обнаружено и прямо противоположное явление: корреляция становится достоверно положительной у матерей на зонах Хартмана и Карри, обладающих повреждающим действием, однако эти матери одновременно формировались также и на благоприятных зонах, которые подпитывали их энергией. Таким образом, почвенные условия и геоактивные зоны Земли являются мощными факторами, влияющими на наследуемость продуктивности у сосны обыкновенной, и их следует использовать в лесной селекции. Для проверки влияния геоактивных зон на рост потомства необходимы тест-культуры и изучение их в течение 5–7 лет.
Работа выполнена при частичной финансовой поддержке задания 2014/153 государственных работ в сфере научной деятельности в рамках базовой части госзадания Минобрнауки России, проект 144 № ГР 01201461915.
Библиографическая ссылка
Рогозин М.В. ПРИЧИНЫ ОТРИЦАТЕЛЬНОЙ НАСЛЕДУЕМОСТИ У СОСНЫ ОБЫКНОВЕННОЙ // Международный журнал прикладных и фундаментальных исследований. 2016. № 8-5. С. 755-759;URL: https://applied-research.ru/ru/article/view?id=10163 (дата обращения: 01.07.2026).