

В условиях Северо-Запада России основными лесообразующими видами являются сосна обыкновенная (Pinus sylvestris L.) и береза повислая (Betula pendula Roth). Ранее нами было проведено сравнительное исследование закономерностей варьирования переменных водного режима этих видов при их совместном произрастании в условиях достаточной почвенной влагообеспеченности [4]. В результате были получены косвенные свидетельства более высокой проводимости ксилемы березы по сравнению с сосной и выявлены отличия в величинах скорости водного потока по ксилеме ствола. Эти отличия были обусловлены, прежде всего, различиями в строении водопроводящих систем хвойного и лиственного видов. Анализ величин дневных и предрассветных водных потенциалов охвоенных побегов указал на наличие у деревьев сосны постоянного водного дефицита, который оказывал значительное влияние на фотосинтез [2]. Данное исследование посвящено оценке величины водного дефицита у растений березы повислой и его влияния на динамику фотосинтеза в условиях достаточной почвенной влагообеспеченности.
Материалы и методы исследования
Исследования проводили в сосняке черничном свежем европейской части средней тайги в Южной Карелии (N 62 °13′, Е 34 °10′). Таксационное описание древостоя и характеристика почв представлены в работе [1]. Объектами исследования послужили 20-летние деревья березы повислой (Betula pendula Roth). Сопряженные исследования водных потенциалов облиственных побегов и фотосинтеза проводили с мая по сентябрь в течение вегетационных периодов 1977-1997 и 2013 гг. Водные потенциалы облиственных побегов (Ψ) определяли с помощью камеры давления Plant Moisture Vessel SKPM 1400 (Skye Instruments Ltd., Великобритания). Регистрацию СО2-газообмена облиственных побегов березы проводили с помощью многоканальной автоматической установки на базе стационарного инфракрасного газоанализатора Infralyt-4 (VEB Junkalor Dessau, Германия) и портативной фотосинтетической системы Li-Cor 6400ХТ (Li-Cor Inc., США). Водные потенциалы почвы измеряли с помощью тензиометра [5]. Влажность почвы определяли термовесовым методом. Образцы почвы отбирали по генетическим горизонтам в верхней части профиля и до глубины 50 см последовательно через каждые 10 см. Метеорологические параметры определяли стандартными метеоприборами и с помощью системы Li-Cor 6400ХТ (Li-Cor Inc., США). Для обработки экспериментальных данных использовали методы вариационной статистики. Проверку гипотез и оценку существенных различий между средними величинами осуществляли с помощью критерия Стьюдента при 5 %-ном уровне значимости.
Результаты исследования и их обсуждение
Анализ многолетних данных суточной динамики водных потенциалов облиственных побегов (Ψ) березы выявил постоянство диапазонов их предрассветных (Ψ.max) и дневных (Ψmin) значений, которые составили – 0.1…– 0.45 и – 0.5…– 1.6 МПа соответственно. Это позволило рассчитать средние за вегетацию величины Ψmax и Ψmin, равные соответственно – 0.24 ± 0.01 и – 1.02 ± 0.03 МПа. При сравнении максимальных и минимальных значений Ψ в разные годы исследования значимых различий не обнаружено (р < 0.05), что свидетельствует о постоянстве показателей в межгодовой динамике.
Наблюдаемые нами в течение суток и вегетационных периодов изменения Ψ были связаны, прежде всего, с изменением погодных условий, поскольку запасы влаги в почве сосняка черничного свежего были достаточно велики. Водные потенциалы почвы в сосняке черничном свежем имели высокие значения в течение всех лет исследований. Минимальные значения, наблюдаемые в наиболее сухие периоды лета (конец июля – начало августа), не опускались ниже – 0.05 МПа. Хорошая увлажненность почвы была обусловлена наличием довольно мощного пласта грунтовых вод, уровень которых изменялся от 1.0 до 1.5 м, и количеством атмосферных осадков в период с мая по сентябрь, которые превышали климатическую норму (342 мм) и составляли в разные годы исследований 350–390 мм.
Сопряженное исследование предрассветных величин Ψmax и запасов влаги в 0–50 мм слое почвы в зоне наибольшего распространения корней деревьев не выявило зависимости между этими показателями. Полученное «несоответствие» между влагообеспеченностью почвы и Ψmax в условиях достаточного почвенного увлажнения обусловлено, вероятно, недостаточно продолжительным темновым периодом северного лета для восстановления водного потенциала растений и установления равновесия показателя в системе «почва – растение». Наряду с этим, несмотря на высокий уровень запасов влаги в почве, полученный результат указывает на наличие водного дефицита в самих растениях. При этом формирование водного дефицита в растениях происходило в основном в дневное послеполуденное время, когда транспирационные расходы влаги не восполнялись приходом ее из почвы.
Наличие постоянного водного дефицита в растениях оказывало влияние на фотосинтез (Р) березы. Синхронная регистрация суточной динамики Р и Ψ березы показала, что от уровня водного дефицита, сформировавшегося к предрассветному часу, в определенной мере зависело время наступления максимума фотосинтеза (Рmax) и его величина. Наиболее высокий водный дефицит, на что указывали величины Ψmax, равные – 0.35…– 0.45 МПа, отмечали, прежде всего, в июне в период «белых ночей», когда небольшой транспирационный расход влаги происходил практически всю ночь. Кроме того, такие значения Ψmax наблюдали в периоды «атмосферных засух», которые чаще всего приходились на конец июля – начало августа [3]. В такие периоды время наступления максимального фотосинтеза сдвигалось на более ранние утренние часы, и депрессия фотосинтеза продолжалась более длительное время. Это приводило к снижению среднедневной продуктивности фотосинтеза.
Дальнейший анализ суточных кривых Р и Ψ облиственных побегов березы показал, что не только время наступления Рmax, но и его величина во многом зависит от уровня водного дефицита в облиственных побегах (рисунок).
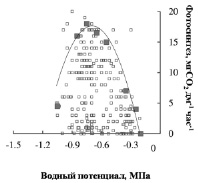
Зависимость интенсивности фотосинтеза от водного потенциала побегов березы
На рисунке представлены результаты одновременной регистрации Ψ и Р, полученные в течение одного вегетационного периода (май – сентябрь). Реализация максимального за сутки фотосинтеза (Рmax) происходила в определенном диапазоне Ψ, который не зависел от года наблюдений. При этом для березы этом диапазон был довольно узким и в среднем составил – 0.7 ± 0.1 МПа. Полученная величина была выше среднего за вегетацию Ψmin. Напротив, у деревьев сосны он был достаточно широким (– 0.7…– 1.1 МПа) и его среднее значение (– 0.9 ± 0.1 МПа) было близко к среднему за вегетацию значению Ψmin ср. [2]. Полученные результаты свидетельствуют, прежде всего, о более высокой требовательности березы к ее влагообеспеченности, по сравнению с сосной.
Поскольку взаимосвязь водного режима и фотосинтеза осуществляется, прежде всего, через устьичную регуляцию, можно предположить, что выявленная величина Ψ – это значения, при которых у березы происходит частичное закрытие устьиц днем, т.е. когда «гидравлические соотношения преобладают над фотоактивными реакциями» [9]. Отмечается также видоспецифичность «пороговых» значений Ψ для древесных растений [9]. При этом высокая чувствительность устьиц к снижению гидравлической проводимости была отмечена как для лиственных [7], так и для хвойных древесных видов [10]. Уменьшение проводимости ксилемы и связанное с ним снижение проводимости устьиц приводит к уменьшению фотосинтеза. Продуктивность растений также связывают с их гидравлической структурой, которая, качественно и количественно определяя способность растений проводить воду от корней к листьям, контролирует максимальную проводимость устьиц и, таким образом, влияет на фотосинтез. Так, было показано, что с увеличением размера дерева общая гидравлическая проводимость снижается [6], вызывая более раннее закрытие устьиц для уменьшения потерь воды и предотвращения развития повреждающих градиентов водного потенциала, что, в свою очередь, приводит к снижению межклеточной концентрации CO2 и интенсивности фотосинтеза [8].
Заключение
В условиях достаточного почвенного увлажнения нами выявлены диапазоны изменчивости предрассветных и дневных водных потенциалов облиственных побегов растений березы повислой и установлена относительная стабилизация их в межгодовой динамике. Нами показано, что одним из внутренних параметров дерева, оказывающих существенное влияние на динамику фотосинтеза, является его водный статус. Даже в условиях достаточного почвенного увлажнения в стволах, ветвях и побегах березы формируется водный дефицит, значительный в отдельные периоды вегетации. Это связано с тем, что гидравлическая архитектура древесных растений, определяющая способность растений проводить воду от почвы к местам использования, контролирует максимальную проводимость устьиц и, таким образом, влияет на фотосинтез.
Работа выполнена в рамках государственного задания ИЛ КарНЦ РАН (проект № 0220-2014-0001).
Библиографическая ссылка
Сазонова Т.А., Болондинский В.К., Придача В.Б., Новичонок Е.В. Влияние водного дефицита листа на фотосинтез березы повислой // Международный журнал прикладных и фундаментальных исследований. 2016. № 10-4. С. 595-597;URL: https://applied-research.ru/ru/article/view?id=10396 (дата обращения: 02.07.2026).