

Аномалии развития желчных пузырей у различных млекопитающих и человека начали описывать более 200 лет тому назад. На рисунке приведены обобщающие данные о различных вариантах аномально развившихся желчных пузырей [1]. Согласно этим данным, раcщепленный (разделенный) желчный пузырь, состоит из двух органов с общим выводным протоком (рисунок, С–F), а двойной желчный пузырь имеет два органа с самостоятельными пузырными протоками, в дальнейшем образующими общую структуру, сливающуюся с общим печеночным протоком (рисунок, G).
Нарушения эмбрионального развития, приводящие к появлению различных вариантов аномальных желчных пузырей [2], обнаружены как у домашних животных, так и человека. Так, частота данной патологии у кошек составляет 1:8, телят 1:28, овец 1:85, свиней 1:198 [1]. У человека же наличие различных патологий желчного пузыря (аномалии количества, размеров, положения, формы) занимает 6–8 % патологий развития всех органов [3], но при этом частота ращепления (неполного удвоения) или полного удвоения желчного пузыря по различным данным составляет 0,02–0,03 % [1–6].
В этих работах приведены сведения о частоте возникновения нарушения процессов органогенеза, различных вариантах анатомической структуры аномальных желчных пузырей. Однако гистологический анализ их проводился крайне редко и в ограниченном объеме.
Объектом, на которых проводились наши исследования, были морские свинки. При вскрытии 184 интактных самцов у одного из них была обнаружена патология желчного пузыря. В доступной нам литературе не было найдено никаких сведений об аномалиях развития желчного пузыря морской свинки. Цель нашего исследования – морфологическое изучение аномального желчного пузыря морской свинки с обращением особого внимания на особенности пролиферативных процессов клеток эпителиальной выстилки.
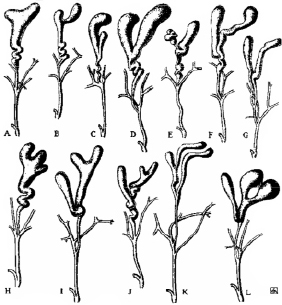
Варианты анормальных желчных пузырей кошки, сформированных в результате нарушения эмбрионального развития: C–F – «раcщепленные» желчные пузыри; G–Y – образный «двойной» желчный пузырь [1]
Материалы и методы исследования
Исследование было проведено на аномальном желчном пузыре, взятом от половозрелого самца морской свинки, массой тела 250 грамм, полученном из питомника «Рапполово». Для того, чтобы избежать методических ошибок при подсчете пролиферативных процессов и сравнении их с литературными данными забой животных (16 часов 15 марта) производился с учетом особенностей циркадных ритмов митотически делящихся и ДНК синтезирующих клеток [7]. За 1 час до забоя всем животным, в том числе и самцу с обнаруженной в дальнейшем патологией развития, вводили Н-3 тимидин (удельная активность15 кюри /ммоль из расчета 0,5 ммкюри/г массы тела). Орган фиксировали в жидкости Буэна, заливали в парафин, депарафинизированные срезы толщиной 7 мкм покрывали эмульсией, экспонировали. Препараты окрашивали гематоксилин-эозином, азаном по Генденгайну, Мак-Манусу, Хейлу. Подсчет ДНК синтезирующих и митотически делящихся клеток производили на 3 тысячи клеток. На этих препаратах изучали строение органа, его стенки, особенности эпителиальной выстилки.
Результаты исследования и их обсуждение
Макроскопический анализ показал, что аномальный желчный пузырь морской свинки является двойным, т.е. представлен двумя желчными пузырями, каждый из которых состоит из дна, тела и шейки органа. Стенка желчного пузыря представлена тремя оболочками: слизистой, мышечной и соединительнотканной.
Слизистая оболочка каждого из двойного (аномального) желчного пузыря образует складки, основу которых составляют прослойки рыхлой соединительной ткани, кровеносных и лимфатических сосудов и нервов. Количество и характер складок, как известно, зависит от степени наполнения органа. Размеры как одного (левого), так и другого (правого) двойного (аномально) желчного пузыря морской свинки не отличались друг от друга. Собственный слой слизистой оболочки представлен рыхлой соединительной тканью. Обращают на себя внимание кровеносные сосуды, имеющие тонкую стенку венозного типа. Во многих случаях сосуды эти располагаются в середине складки, способствуя, по-видимому, сохранению ее месторасположения. Под эпителием располагаются мелкие кровеносные сосуды капиллярного типа, образующие густую сеть – «plexus subepithelialis». Клеточные элементы собственного слоя представлены фибробластами, клетками типа ретикулярных и адвентициальных. Вытянутой веретенообразной формы фибробласты располагаются непосредственно под базальной мембраной. Подсчет количества эпителиальных клеток и расположенных под ним уплощенных фибробластов показал, что один фибробласт принимает участие в образовании базальной мембраны для 4,5 эпителиальных клеток. Собственный слой слизистой как одного, так и другого двойного желчного пузыря не отличается как друг от друга, так и от нормально сформированного органа. Гистологический анализ t. fibromuscularis показал, что пучки гладкомышечных клеток, идущих в разных направлениях, перемежаются весьма незначительными прослойками соединительной ткани. Перимускулярный слой состоит из пучков коллагеновых волокон и клеток, преимущественно фибробластического ряда. В этом слое располагаются артерии, вены, лимфатические сосуды. Со стороны брюшной полости желчные пузыри покрыты серозной оболочкой, мезотелий которой представлен уплощенными клетками, среди которых обнаружены и двухъядерные. В каждом из двойных желчных пузырей морской свинки собственный слой слизистой оболочки, фибромускулярный, перимускулярный, а также серозная оболочка ничем по своим характеристикам не отличаются как друг от друга, так и от нормально развившихся желчных пузырей.
Эпителиальная выстилка аномального желчного пузыря морской свинки представлена однослойным однорядным высокопризматическим эпителием. На поверхности эпителиального пласта тонкая каемка дает реакцию на кислые и нейтральные мукополисахариды. У основания складок эпителиальные клетки принимают подчас изопризматическую форму. Основная масса клеток, представлена высокопризматическим эпителием. Небольшое количество сравнительно широких «бочкообразных» клеток, как правило, располагаются попарно и, вероятно, как было высказано нами ранее, являются дочерними клетками после только что закончившегося деления. Клетки эти располагаются у основания складок, т.е. тех местах, где обнаруживается основное количество пролиферирующих клеток. Третью немногочисленную группу образуют так называемые «штифтиковые» (Stiftzellen) клетки, разбросанные поодиночке или сгруппированные по 2–3, имеющие узкую интенсивную окрашенную цитоплазму – по Мак-Манусу красным цветом, а по Хейлу голубым, что свидетельствует о наличии как кислых, так и нейтральных мукополисахаридов. Однако из апикальной части этих клеток в просвет желчного пузыря слизь не выделяется. Сравнительный анализ полученных данных с литературными [6] свидетельствует о том, что в ядрах таких клеток не было обнаружено включений Н-3 тимидина. Все вышеописанные формы структур эпителиального пласта, по нашему мнению, вероятно, находятся на разных этапах жизненного цикла эпителиальных клеток желчного пузыря. Кроме эпителиальных клеток, вырабатывающих слизь в небольших количествах, в составе покровного пласта аномальных желчных пузырей морской свинки выявлены и единичные (1 %) специализированные железистые элементы – бокаловидные клетки. В этой же области, т.е. шейке желчного пузыря, у аномальных животных расположены единичные слизистые железы. В других частях аномального желчного пузыря слизистые железы не были обнаружены.
Сравнительная характеристика морфологических особенностей аномально развившегося (двойного) желчного пузыря по сравнению с интактными одиночными производилась с учетом сведений о патологических процессах в желчных пузырях, описанных в руководстве по патологии [8]. Гистологический анализ показал отсутствие как специфических для данного органа (наличие в составе эпителиального пласта лишь незначительного числа бокаловидных клеток (1 %), единичных слизистых желез, количество которых резко увеличивается при патологии желчного пузыря), так и неспецифических (отсутствие классических признаков воспаления и других морфологических особенностей, присущих поврежденным тканям) показателей, свидетельствующих об отсутствии патологических состояний в исследованном нами двойном желчном пузыре.
В следующей части исследования приведены результаты, характеризующие пролиферативную активность эпителиальных клеток различных частей аномальных желчных пузырей морской свинки. Анализ табл. 1 свидетельствует о том, что митотическая активность клеток эпителия каждого из аномальных желчных пузырей морской свинки колеблется в незначительных пределах как в разных отделах, так и в органе в целом.
Анализ табл. 2 свидетельствует о том, что количество ДНК синтезирующих клеток эпителия в аномальных желчных пузырях варьирует в незначительных пределах.
Таблица 1
Показатели митотической активности (МК в %о) клеток эпителиального пласта, выстилающих разные части нормальных и двойного (аномального) желчных пузырей морской свинки (M ± m)1
|
Исследуемый участок эпителиальной выстилки |
Митотическая активность клеток эпителия (в %о) |
||
|
нормального желчного пузыря* |
аномального (двойного) желчного пузыря** |
||
|
1*** |
2*** |
||
|
дно желчного пузыря |
1,3 ± 0,4 |
1,7 |
2,0 |
|
тело желчного пузыря |
1,6 ± 0,5 |
2,0 |
2,3 |
|
шейка желчного пузыря |
2,2 ± 0,5 |
2,3 |
2,7 |
|
весь желчный пузырь |
1,7 ± 0,4 |
2,0 |
2,3 |
Примечания: 1M ± m – среднее арифметическое ± ошибка среднего; * – литературные данные [9]; ** – результаты данного исследования; *** – двойной желчный пузырь: 1) левый, 2) правый.
Таблица 2
Количество ДНК синтезирующих (ИМЯ в %о) клеток эпителиального пласта, выстилающих разные части нормальных и двойного (аномального) желчных пузырей морской свинки (M ± m)1
|
Исследуемый участок эпителиальной выстилки |
Количество ДНК синтезирующих клеток (в %о) |
||
|
нормального желчного пузыря* |
аномального (двойного) желчного пузыря** |
||
|
1*** |
2*** |
||
|
дно желчного пузыря |
6,2 ± 0,6 |
5,3 |
7,0 |
|
тело желчного пузыря |
7,0 ± 0,7 |
6,0 |
7,3 |
|
шейка желчного пузыря |
7,2 ± 0,7 |
6,7 |
7,3 |
|
весь желчный пузырь |
6,7 ± 0,5 |
6,0 |
7,1 |
Примечания: 1M ± m – среднее арифметическое ± ошибка среднего; * – собственные (ранее не опубликованные) данные; ** – результаты данного исследования; *** – двойной желчный пузырь: 1) левый, 2) правый.
Таблица 3
Топография пролиферирующих эпителиальных клеток нормального и двойного (аномального) желчных пузырей морской свинки в различных участках эпителиальной выстилки (M ± m)1
|
Исследуемый участок эпителиальной выстилки |
Количество в изучаемых участках эпителиальной выстилки (в % от общего числа пролиферирующих клеток) |
|||||
|
Митозов |
ДНК синтезирующих клеток |
|||||
|
эпителия нормального желчного пузыря* |
эпителия аномального (двойного) желчного пузыря** |
эпителия нормального желчного пузыря* |
эпителия аномального (двойного) желчного пузыря** |
|||
|
1*** |
2*** |
1*** |
2*** |
|||
|
Вершина складок |
6,0 ± 1,2 |
7,3 |
6,7 |
2,0 ± 0,5 |
2,3 |
2,7 |
|
Боковая часть |
21,0 ± 2,0 |
23,0 |
22,0 |
10,0 ± 1,1 |
8,0 |
10,3 |
|
Основание складок |
73,0 ± 2,2 |
70,7 |
71,3 |
88,0 ± 1,1 |
90,7 |
87,0 |
Примечания: 1M ± m – среднее арифметическое ± ошибка среднего; * – литературные данные [9]; ** – результаты данного исследования; *** – двойной желчный пузырь: 1) левый, 2) правый.
В табл. 3 представлены результаты нашего исследования в сопоставлении с литературными данными [9] о топографии ДНК синтезирующих и митотически делящихся клеток в различных участках эпителиального пласта, выстилающего различные участки складок желчного пузыря морских свинок. Ранее было показано, что подавляющее большинство пролиферирующих клеток призматического эпителия нормального желчного пузыря морских свинок находится у основания складок и лишь незначительная часть на их вершине, что характеризует наличие зоны камбиальных клеток, сходной по своей топографии с клетками эпителия, выстилающими крипты кишечника [9]. Полученные нами данные о том, что синтезирующие ДНК и митотически делящиеся клетки эпителия двойного желчного пузыря находятся у основания складок, как и в нормальном органе (табл. 3), свидетельствуют о том, что и в аномально развившемся пузыре камбиальные зоны, а также, по-видимому, и скорость перемещения эпителиального пласта оказались неизмененными.
В заключение необходимо отметить, что работа с 184 морскими свинками выявила одну особь с аномально развившимся (двойным) желчным пузырем. Морфологическими методами было показано, что каждый из двух желчных пузырей состоял из дна, тела и шейки, стенки которых по своему гистологическому строению соответствовали нормальному. Эпителиальная выстилка каждого изученного нами пузыря была представлена типичными для данного эпителия клетками, в том числе и 1 % высокоспециализированными клетками – бокаловидными, а также единичными (находящимися только в шейке органа) слизистыми железами. Признаков патологических процессов в аномальных желчных пузырях обнаружено не было. Подсчет количества митотически делящихся и ДНК синтезирующих клеток эпителия, их распределения позволил получить уникальные первые данные о пролиферативных процессах в двойном желчном пузыре и наличии камбиальных зон у основания складок. Несмотря на то, что выборка единична и не может быть подвергнута статанализу, полученные результаты представляют большой интерес, ибо являются пионерскими.
Библиографическая ссылка
Дыбан П.А. ДВОЙНОЙ ЖЕЛЧНЫЙ ПУЗЫРЬ МОРСКОЙ СВИНКИ: МОРФОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА И ПРОЛИФЕРАТИВНАЯ АКТИВНОСТЬ // Международный журнал прикладных и фундаментальных исследований. 2017. № 11-2. С. 233-237;URL: https://applied-research.ru/ru/article/view?id=12005 (дата обращения: 30.06.2026).