

Этническая группа, представляющая жителей Северной Осетии, является в какой-то степени уникальной. Достаточно долгое время ведется множество дискуссий по поводу ее происхождения и о том, могут ли осетины Северного Кавказа претендовать на статус прямых потомков алан и Аланского царства [1]. Аланы в свою очередь представляли собой ираноязычное кочевое племя скифо-сарматского происхождения, в письменных источниках упоминаются с 1 века нашей эры, времени их появления в Приазовье и Предкавказье. Учитывая все это, осетины могут являться уникальным и единственным сохранившимся реликтом североиранского (скифо-сарматского) языкового мира [2]. На сегодняшний день есть данные об анализе митохондриальной ДНК и Y-гаплогрупп у народов Северной и Южной Осетии, позволяющие строить предположения относительно вероятного происхождения осетин и их сходства или различия с другими популяциями [3]. Однако данных по частотам встречаемости генов гистосовместимости HLA, которые так же могли бы послужить источником для подобного рода размышлений, нет. Кроме того, в свете последних тенденций по расширению российских национальных регистров потенциальных доноров костного мозга требуется включение в них как можно большего количества человек различных национальностей. Исходя из всего вышесказанного, типирование такой уникальной народности, как осетины, по генам гистосовместимости HLA представляется особенно интересным.
Цель данной работы – оценить частоты встречаемости генов HLA-A, -B, -C, -DRB1 и -DQB1 у осетин Северного Кавказа.
Материалы и методы исследования
Для исследования были привлечены потенциальные доноры костного мозга регистра ФГБУ ГНЦ МЗ РФ, рекрутированные в период с октября по ноябрь 2016 года. Исследуемая выборка представлена 120 здоровыми донорами. Все доноры проживают в г. Владикавказе Северной Осетии, являются осетинами в третьем поколении. В качестве материала для исследования использовали венозную кровь, взятую в пробирки с 0,5 % раствором ЭДТА. Выделение ДНК проводили на приборе NorDiag Arrow с использованием соответствующих наборов Blood DNA 200 и 500 Extraction Kit (NorDiag, Норвегия). Типирование по 5 локусам генов HLA проводили методом PCR-SSO на платформе Luminex с использованием наборов Lifecodes HLA-A/B/C/DRB1/DQA1, DQB1 SSO Typing Kit (Immucor, США). Результаты типирования по локусу HLA-DQA1 в данной работе не представлены. Статистическую обработку данных проводили с использованием программы Arlequin 3.5 для оценки частот аллелей методом максимального правдоподобия с помощью алгоритма максимизации ожидания для данных. Соответствие наблюдаемого распределения аллельных вариантов равновесию Харди – Вайнберга так же определялось в компьютерной программе Arlequin 3.5. Филогенетическая дендрограмма была создана с применением программы PHYLIP, на основе частот аллелей и подсчета генетических расстояний, вычисленных по Нею [4].
Результаты исследования и их обсуждение
В ходе данного исследования в локусах HLA-А, -В и -С в популяции осетин было выявлено 14, 19 и 13 аллельных групп соответственно. В локусах HLA-DRB1 и –DQB1 выявлено 12 и 5 аллельных групп. Согласно закону Харди – Вайнберга популяция доноров Северной Осетии находится в равновесии по всем локусам, кроме HLA-DQB1. Возможно, это связано с немногочисленной выборкой в нашем исследовании.
Наиболее распространенными аллелями у осетин в локусе А оказались А*02 (частота встречаемости 0,238), *30 (0,129), *01 (0,121) и *03 (0,117), *24 (0,1). Большинство наблюдаемых значений частот в локусе А у осетин представляет собой промежуточные значения между европейскими и азиатскими популяциями. Так, высокая частота аллеля А*02 является признаком типично европейской популяции, в то время как высокая частота А*30 встречается в основном среди азиатов (в том числе у иранских народов) и у представителей африканского континента. Однако в некоторых европейских популяциях так же имеются схожие с популяцией осетин данные по частоте встречаемости аллеля А*30 (напр., Spain Murcia [5]). В нашем исследовании в выборке осетин не встретились следующие аллели из локуса HLA-A: A*34, *36, *43, *69, *74, *80, не характерные для европейцев.
Частота генов HLA-A, -B, -C, -DRB1, -DQB1 в популяции осетин
|
Аллель |
Количество |
Частота аллеля |
Аллель |
Количество |
Частота аллеля |
Аллель |
Количество |
Частота аллеля |
||
|
HLA-A |
B*38 |
11 |
0,046 |
HLA-DRB1 |
||||||
|
A*01 |
28 |
0,121 |
B*39 |
3 |
0,013 |
DRB1*01 |
12 |
0,050 |
||
|
A*02 |
57 |
0,238 |
B*40 |
10 |
0,042 |
DRB1*03 |
30 |
0,125 |
||
|
A*03 |
28 |
0,117 |
B*44 |
13 |
0,054 |
DRB1*04 |
17 |
0,071 |
||
|
A*11 |
11 |
0,046 |
B*48 |
1 |
0,004 |
DRB1*07 |
30 |
0,125 |
||
|
A*23 |
7 |
0,029 |
B*49 |
39 |
0,163 |
DRB1*08 |
7 |
0,029 |
||
|
A*24 |
24 |
0,100 |
B*50 |
15 |
0,063 |
DRB1*10 |
2 |
0,008 |
||
|
A*25 |
4 |
0,013 |
B*51 |
31 |
0,129 |
DRB1*11 |
47 |
0,196 |
||
|
A*26 |
9 |
0,038 |
B*55 |
1 |
0,004 |
DRB1*12 |
1 |
0,004 |
||
|
A*30 |
31 |
0,129 |
B*57 |
3 |
0,013 |
DRB1*13 |
52 |
0,217 |
||
|
A*31 |
5 |
0,021 |
HLA-C |
DRB1*14 |
7 |
0,029 |
||||
|
A*32 |
21 |
0,088 |
С*01 |
1 |
0,004 |
DRB1*15 |
30 |
0,125 |
||
|
A*33 |
2 |
0,008 |
С*02 |
14 |
0,058 |
DRB1*16 |
5 |
0,021 |
||
|
A*66 |
2 |
0,008 |
С*03 |
11 |
0,046 |
HLA-DQB |
||||
|
A*68 |
11 |
0,046 |
С*04 |
20 |
0,083 |
DQB1*02 |
56 |
0,233 |
||
|
HLA-B |
С*05 |
5 |
0,021 |
DQB1*03 |
73 |
0,304 |
||||
|
B*07 |
23 |
0,096 |
С*06 |
38 |
0,158 |
DQB1*04 |
4 |
0,017 |
||
|
B*08 |
16 |
0,067 |
С*07 |
101 |
0,421 |
DQB1*05 |
27 |
0,113 |
||
|
B*13 |
18 |
0,075 |
С*08 |
2 |
0,008 |
DQB1*06 |
80 |
0,333 |
||
|
B*14 |
1 |
0,004 |
С*12 |
19 |
0,079 |
|||||
|
B*15 |
11 |
0,046 |
С*14 |
9 |
0,038 |
|||||
|
B*18 |
17 |
0,071 |
С*15 |
7 |
0,029 |
|||||
|
B*27 |
7 |
0,029 |
С*16 |
11 |
0,046 |
|||||
|
B*35 |
17 |
0,071 |
С*17 |
2 |
0,008 |
|||||
|
B*37 |
3 |
0,013 |
||||||||
Примечание. Жирным шрифтом выделены наиболее частотные варианты.
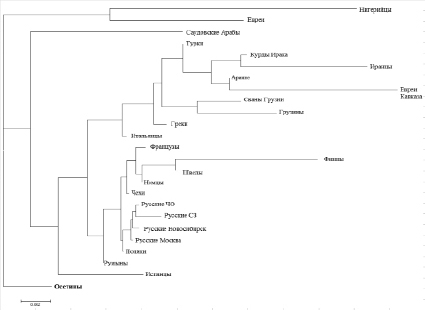
Дендрограмма генетических расстояний, построенная на основе частот генов HLA-A, -B, -DRB1 в популяции осетин и некоторых мировых популяций
В локусе HLA-В наиболее частотным оказался аллельный вариант В*49 (0,154), что является отличительной особенностью осетин (как представителей народов Кавказа), далее следует В*51 (0,129), B*07 (0,096), В*13 (0,075). Аллель В*49 с высокой частотой встречается у израильско-эфиопских евреев (0,187), камерунского народа Бамилеке (0,143, AF), в Руанде (0,116) и Армении (0,095) [5, 6]. То есть B*49 прежде всего характерен для населения регионов Южной Азии и части африканского континента. Повышенные частоты встречаемости В*13 и В*51 также более всего характерны для азиатских популяций. В исследуемой выборке аллели HLA-B*41, *42, *45, *46, *47, *52, *53, *54, *56, *58, *59, *67, *73, *78, *81, *82 не были нами обнаружены.
Локус С у осетин характеризуется преобладанием аллеля HLA-C*07 (частота встречаемости 0,421). Далее по частоте следуют С*06 (0,158) и *04 (0,083). Преимущественный перевес в частоте встречаемости аллеля С*07 в локусе HLA-C характерен для большинства мировых популяций (таблица). Низкочастотный для европейских популяций вариант HLA-C*18 в исследуемой выборке осетин не был выявлен.
При оценке частот встречаемости генов HLA II класса наиболее распространенными среди осетин оказались аллельные варианты HLA-DRB1*13 (0,217) и *11 (0,196), HLA-DQB1*06 (0,333) и *03 (0,304). Довольно высокая частота аллеля DRB1*13 является отличительной чертой осетин, в особенности по сравнению с европейскими популяциями, где данный аллель встречается в полтора раза реже. Ближе всего по частоте встречаемости аллеля DRB1*13 к осетинам оказались арабы Туниса (0,21), сваны Грузии (0,175) [5]. Наиболее редкими вариантами генов HLA II класса среди осетин оказались HLA-DRB1*10, *12 и HLA-DQB1*04. Вариант HLA-DRB1*09 не встретился в нашем исследовании ни разу. Это в общем-то не вызывает удивления, поскольку данный аллельный вариант, характерный для некоторых азиатских популяций, встречается в других мировых популяциях с довольно низкой частотой, и небольшой размер нашей выборки, вероятно, просто не позволил его уловить.
На основе частот аллелей локусов HLA-A, -B и -DRB1 был проведен анализ генетического родства некоторых мировых популяций с исследуемой группой. По данным о генетических расстояниях была построена дендрограмма (рисунок).
В качестве групп сравнения были выбраны следующие популяции: этнические группы России – русские Москвы [7], русские ЧО (Челябинской области) [8], русские Новосибирска [9], русские СЗ (Северо-Запада России) [5], популяции стран Средиземноморья и Юго-Западной Азии – евреи (Israel Ethiopia Jews), саудовские арабы (Saudi Arabia) [5], турки [10], иранцы [11], евреи Кавказа (Israel Kavkazi Jews), сваны Грузии (Georgia Svaneti Region Svan), греки (DKMS Greece minority), итальянцы (DKMS Italian minority), испанцы (Spain Murcia) [5], курды Ирака [12], армяне [6], грузины [2], популяции Европы – поляки [13], немцы (Germany pop6), шведы (Swedes), финны (Finnes), французы (French pilot study on Bone Marrow Registry), румыны (DKMS Romania minority) и нигерийцы (Nigerians) [5].
Как видно на представленной дендрограмме, популяция осетин имеет довольно обособленное положение не только относительно других популяций Российской Федерации, но и относительно остальных мировых популяций, что разумеется связано как с особенностями территориального проживания, так и этногенеза исследуемой группы. Наименьшее генетическое расстояние из рассматриваемых популяций осетины показывают с итальянцами (генетическое расстояние по Нею 1,08), румынами (1,12) и чехами (1,2). Из популяций Юго-Западной Азии ближе всего осетины оказались к туркам (1,23). Наибольшие значения генетических расстояний осетины имеют с финнами (3,26), евреями Кавказа (3,31) и нигерийцами (3,07). Осетины оказались весьма удалены от других кавказских популяций, включенных в сравнение (армян, грузин, сванов, евреев Кавказа).
Заключение
Таким образом, популяция осетин действительно представляет собой уникальную этническую группу, сочетающую в себе черты как азиатских, так и европейских популяций. Об этом свидетельствует наличие в генотипах осетин одновременно таких аллельных вариантов, как А*02 и А*30, В*07 и B*51, B*49, которые встречаются у них с довольно высокой частотой. Кроме того, популяция осетин обладает своими индивидуальными, присущими только ей чертами. К сожалению, в доступных источниках нет данных о распределении частот генов HLA у соседних по географическому положению популяций, таких как ингуши, карачаевцы, черкесы, кабардинцы. Поэтому на сегодняшний день трудно судить, можно ли выявленные особенности считать уникальными для осетин или они характерны и для других народов Северного Кавказа. Так, отличительной чертой осетин является высокая частота аллелей B*49 и DRB1*13, выделяющая ее среди остальных этнических групп Юго-Западной Азии и Кавказа. Возможно, именно эти особенности являются результатом влияния на становление иммуногенетического профиля осетин Северного Кавказа древнего племени алан. Данные о частотах встречаемости аллелей генов HLA среди осетин могут стать интересной базой для дальнейшего исследования этногенеза популяций Северного Кавказа, кроме того могут быть использованы для проведения научных исследований в области связи генов системы HLA c предрасположенностью к различным заболеваниям. Обладая своими индивидуальными особенностями популяция осетин представляет собой уникальный материал для включения в региональный регистр потенциальных доноров костного мозга, поскольку наличие в регистре представителей с такими уникальными генотипами повышает шанс нахождения неродственного донора для пациентов с редкими HLA-генотипами. Несомненно, требуются дальнейшие исследования в этой области для расширения знаний об иммуногенетическом профиле как популяции осетин, так и других народностей, проживающих на территории Российской Федерации.
Библиографическая ссылка
Кузьминова Е.П., Чапова Р.С., Хамаганова Е.Г. ИММУНОГЕНЕТИЧЕСКИЙ ПРОФИЛЬ (HLA-A, -B, -C, -DRB1, -DQB1) ПОПУЛЯЦИИ ОСЕТИН СЕВЕРНОГО КАВКАЗА // Международный журнал прикладных и фундаментальных исследований. 2017. № 12-2. С. 287-291;URL: https://applied-research.ru/ru/article/view?id=12036 (дата обращения: 01.07.2026).