

Потребность в устойчивых и адаптированных сортах растений для включения их в селекционные программы предопределила необходимость изучения ответных реакций растений на воздействие неблагоприятных условий среды у разных видов и сортов растений. Адаптивный характер реакции растения на стресс сопровождается повышением или понижением содержания тех или иных компонентов клетки, а также развитием любого признака, который способствует выживанию вида и его размножению.
Известно, что семена, которые лучше переносят неблагоприятные условия ускоренного старения, более устойчивы к абиотическим стрессорам при прорастании. Приводятся сведения о связи старения с нарушением физиологических и биохимических процессов [1; 2], с накоплением ингибиторов роста и токсичных продуктов метаболизма [3]. В основе повреждений, приводящих к старению семян и снижению их стресс-устойчивости, лежит генерация свободных радикалов, которую инициирует автоокисление липидов и неферментативное гликозилирование [4]. Эти реакции не требуют высокого влагосодержания, и их продукты способны накапливаться в воздушно-сухих семенах, что приводит к окислительным повреждениям мембран, белков и ДНК. По современным представлениям некоторые аминокислоты, в том числе гетероциклическая аминокислота пролин, инициируют активацию адаптивных механизмов [5; 6]. Пролин уменьшает осмотический стресс, участвует в передаче сигнала стресса, регулирует окислительно-восстановительный потенциал клетки. В условиях стресса аккумуляция пролина индуцируется усилением его синтеза, а также восстановлением окисленных аминокислот. Значительная часть пролина, накапливающегося в клетке во время стресса, синтезируется из глутамата в пероксисомах [5]. При едином мнении, что увеличение содержания пролина в растениях является общей физиологической реакцией на засуху, засоление и другие неблагоприятные воздействия, в литературе встречаются противоречивые сведения о зависимости устойчивости растений к стрессу от содержания пролина. Так, в работе С.И. Михальской и др. проанализирован уровень свободного L-пролина в побегах и корнях проростков кукурузы в условиях сульфатно-хлоридного засоления. Показано, что стресс-устойчивость сопровождается повышением содержания этого осмолита. Солеустойчивость трансформантов, содержащих двухцепочечный РНК-супрессор гена пролиндегидрогеназы кукурузы, сопровождался аккумуляцией свободного L-пролина [7]. В работе Y. Luo и др. более высокое содержание пролина наблюдалось у холодоустойчивого генотипа земляники по сравнению с неустойчивым [8]. А.В. Поморцев также отмечает более высокое содержание пролина у морозоустойчивых растений ржи и тритикале [9]. Сообщается, что существует положительная корреляция между содержанием пролина и устойчивостью к засухе у сортов риса и устойчивостью к засолению у растений табака и картофеля [10].
Согласно противоположной точке зрения, чем хуже растение переносит неблагоприятные условия, тем выше содержание пролина в его тканях [11]. Так, в работе A. Aghaee и др. [12] приводятся данные о более низком содержании пролина в стрессовых ситуациях у устойчивого генотипа риса. По данным J. Liu [13] y чувствительных к засолению и к холоду мутантов арабидопсиса отмечено более высокое содержание пролина. По утверждению А.Ф. Кириллова увеличение содержания пролина может служить в качестве количественной меры водного стресса [11].
Эта противоречивость может быть обусловлена как методическими причинами (различная доза и экспозиция стрессовых воздействий в различных экспериментах), так и сложным механизмом взаимодействия пролина с другими стресс-протекторами. Сравнительное исследование стрессового накопления пролина в различных органах растений, отличающихся по устойчивости видов и сортов необходимо как для разработки методологии оценки устойчивости растений, так и для более глубокого понимания стресс-протекторных функций пролина.
Цель исследования заключалась в сравнительной оценке устойчивости образцов мягкой пшеницы к различным стресс факторам по показателям жизнеспособности семян и по содержанию гетероциклической аминокислоты пролин (пирролидин-α-карбоновая кислота) в листьях растений пшеницы.
Материалы и методы исследования
Объектами исследования служили свежие семена 5 разновидностей мягкой пшеницы: YBFS017 K-31 T. aestivum L. v. erythrospermum Körn., YBFS 017 K-35 T. aestivum L. v. erythroleucon Körn., YBFS017 K-71 T. aestivum L. v. murinum Flaks., YBFS017 K-72 T. aestivum L. v. cainotrics Körn., YBFS 017 K-75 T. aestivum L. v. glaucolutescens Vatr. Стресс-факторами являлись ускоренное старение семян, обезвоживание и засоление проростков. Для имитации продолжительности хранения семян применялся метод их искусственного состаривания. Этот метод предполагает 3-дневную инкубацию семян при повышенной относительной влажности и температуре воздуха [1], что позволяет моделировать воздействие неблагоприятных факторов и прогнозировать их влияние на устойчивость семян различных сортов и образцов растений. Оценка жизнеспособности проводилась по тесту лабораторной всхожести семян, выражаемой в процентах от общего числа (n):
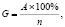
где А – число взошедших семян.
При засухе (содержание проростков в растворе сахарозы (10,5 атм) в течение суток) и солевом стрессе (содержание проростков в растворе 10 атм NaCl (10 атм) в течение суток) анализировали динамику изменения содержания пролина в листьях растений по методу Bates [14]. Стрессовому воздействию подвергали 5-дневные проростки, выращенные в нормальных условиях. Для сравнительного анализа использовали 6-суточные проростки. Пробы отбирали и фиксировали в одно и то же время суток. Контролем служили растения, выращенные в нормальных условиях.
Результаты исследования и их обсуждение
Для оценки функциональных нарушений жизнеспособности семян при ускоренном старении нами использовался такой интегральный показатель, как их всхожесть (рис. 1). При оптимальных условиях прорастания всхожесть семян 5 разновидностей мягкой пшеницы варьировала в пределах 92,5–100,0 %. Анализ всхожести подвергнутых ускоренному старению семян 5 разновидностей мягкой пшеницы показал, что 3-дневное состаривание подавляло прорастание семян различных образцов в разной степени. Так, при наблюдаемом резком падении (на 12,0–46,5 %) всхожести семян у образцов K-71 – T. aestivum L. v. murinum Flaks., K-72 – T. aestivum L. v. cainotrics Körn. и K – 75 T. aestivum L. v. glaucolutescens Vatr., для образцов K-31 – T. aestivum L. v. erythrospermum Körn., K-35 – T. aestivum L. v. erythroleucon Körn. отмечено падение всхожести всего на 2,0 %, что свидетельствует об их большей устойчивости к ускоренному старению. Наибольший процент потери всхожести семян (46,5 %) был характерен для образца YBFS017 K-72 – T. aestivum L. v. cainotrics Körn.
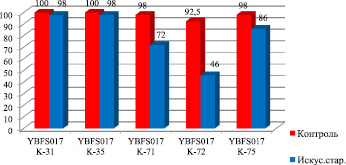
Рис. 1. Всхожесть искусственно состаренных семян различных образцов мягкой пшеницы (YBFS 017 K-31 – T. aestivum L. v. erythrospermum Körn., YBFS 017 K-35 – T. aestivum L. v. erythroleucon Körn., YBFS 017 K-71 – T. aestivum L. v. murinum Flaks., YBFS 017 K-72 – T. aestivum L. v. cainotrics Körn., YBFS 017 K-75 – T. aestivum L. v. glaucolutescens Vatr.)
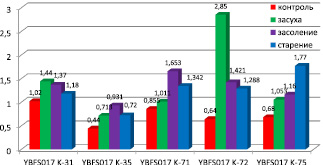
Рис. 2. Содержание пролина у образцов мягкой пшеницы в условиях засухи, засоления и старения (µM/г) (YBFS017 K-31 – T. aestivum L. v. erythrospermum Körn., YBFS 017 K-35 – T. aestivum L. v. erythroleucon Körn., YBFS017 K-71 – T. aestivum L. v. murinum Flaks., YBFS017 K-72 – T. aestivum L. v. cainotrics Körn., YBFS 017 K-75 – T. aestivum L. v. Glaucolutescens Vatr.)
На рис. 2 представлены результаты исследований по определению содержания пролина в побегах проростков, подвергнутых старению, засухе и засолению. Уровень этой аминокислоты у ряда изученных образцов варьировал, однако по абсолютной величине содержание пролина в проростках опытных растений превосходило показатели содержания пролина в проростках контрольных вариантов. Содержание пролина варьировало от 0,44 до 1,02 µM/мг у растений контрольного варианта, где значение содержания пролина имело свой максимум у образца К-31 T. aestivum L. v. erythrospermum Körn., а минимальное значение было отмечено у варианта опыта К-35 T. aestivum L. v. erythroleucon Körn.
Обнаружены значительные различия между образцами по содержанию пролина при стрессе. При ускоренном старении содержание пролина варьировало в пределах 0,72 и 1,77 µM/мг. Наибольшая кратность увеличения содержания пролина отмечена у образца К-72 T. aestivum L. v. сainotrics Körn., в 2 раза выше, чем в контрольном варианте. Относительно контроля в 1,16 раз увеличилось содержание пролина у образца К-31 T. aestivum L. v. еrythrospermum Körn. По мнению Ж.Н. Калацкой и др., так как уровень пролина возрастает пропорционально увеличению степени и продолжительности воздействия неблагоприятных условий хранения, предполагается, что его накопление в проростках свидетельствует скорее о степени воздействия повреждающего фактора, а не о проявлении устойчивости к нему [15]. Однако в наших опытах наблюдалось различное накопление пролина в ответ на одинаковое для всех образцов стрессовое воздействие. Что, опираясь на дополнительные результаты по всхожести семян при ускоренном старении, позволяет нам сделать выводы о детерминированности устойчивости различных генотипов растений.
При засухе диапазон изменения содержания пролина варьировал между 0,713 и 2,85 µM/мг. Отмечено многократное увеличение содержания пролина при засухе относительно контроля в листьях образца К-72 T. aestivum L. v. cainotrics Körn. (в 4,45 раза). Наименьшее изменение содержания пролина относительно контроля наблюдали у разновидности мягкой пщеницы К-71 T. aestivum L. v. murinum Flaks. (в 1,2 раза).
При засолении содержание пролина менялось в пределах 0,931–1,653 µM/мг. Наибольшее значение содержания пролина наблюдалось у растений образца К-71 T. aestivum L. v. murinum Flaks., что превышало значение контроля в 1,9 раз, наименьшее у образца К-35 T. aestivum L. v. erithroleucon Körn., превышение относительно контроля в 2,1 раза. Представляет интерес образец К-72 T. aestivum L. v. cainotrics Körn., у которого содержание пролина превышало значение контрольного варианта в 2,2 раза. Наименьшая кратность превышения содержания пролина относительно контроля была у образца К-31 T. aestivum L. v. erythrospermum Körn. – 1,34 раза.
Интересно, что значительное увеличение содержания пролина в условиях состаривания, засухи и засоления относительно контроля происходило у тех форм, где в контрольных вариантах содержание этой аминокислоты достаточно низкое. В нашем эксперименте таковыми являлись образцы К-35 T. aestivum L. v.еrithroleucon Körn., К-72 T. aestivum L. v. сainotrics Körn. и К-75 T. aestivum L. v. glaucolutescens Vatr., где значения содержания пролина у контрольных растений составляли, соответственно 0,44 µM/мг, 0,64 µM/мг, 0,68 µM/мг.
Заключение
Сравнительная оценка устойчивости семян изученных 5 разновидностей мягкой пшеницы к старению и ответной реакции растений на воздействие засухи и засоления позволила сделать выводы, что по показателям всхожести семян после ускоренного старения и по содержанию пролина в листьях после стрессового воздействия, образец К-31 T. aestivum L. v. erythrospermum Körn. проявил большую устойчивость как к засухе и к засолению, так и к ускоренному старению по сравнению с остальными изученными образцами. В ряду исследованных образцов наименее устойчивым оказался образец К-72 T. aestivum L. v. cainotrics Körn.
Библиографическая ссылка
Мамедова С.А., Ибрагимова З.Ш., Алиев Р.Т. ОЦЕНКА УСТОЙЧИВОСТИ РАЗЛИЧНЫХ ОБРАЗЦОВ ПШЕНИЦЫ К СТАРЕНИЮ, ЗАСУХЕ И ЗАСОЛЕНИЮ // Международный журнал прикладных и фундаментальных исследований. 2018. № 12-1. С. 84-87;URL: https://applied-research.ru/ru/article/view?id=12526 (дата обращения: 11.07.2026).