

Развитие биологических и медицинских подходов к реконструкции зубов с использованием стволовых клеток является перспективным и остается одной из самых серьезных проблем в стоматологической сфере на ближайшие годы [1, 2]. Однако наиболее исследованной моделью для исследования регенерации структур развивающихся зубов являются грызуны, резцы которых непрерывно растут на протяжении всей жизни животного за счет наличия эпителиальных и мезенхимальных стволовых клеток [3, 4]. Исследования по формированию коронкового дентина были одним из основных направлений развития зуба в течение нескольких десятилетий. Несмотря на то, что известно о развитии зубов млекопитающих из двух типов клеток: эктодермы, образующей амелобласты, и клеток эктомезенхимы, которые являются источником одонтобластов и цементобластов, процесс развития зубов и дифференцировки клеток остаётся представленным тупиковыми концепциями. Популяционная распространенность генетической короткой корневой аномалии (SRA) без видимых дефектов коронки близка к 1,3 %. Кроме того, люди с самой SRA предрасположены к корневой резорбции во время ортодонтического лечения [5, 6]. Два типа клеток, дифференцируясь в процессе развития ротовой полости, взаимодействуют и индуцируют весь процесс инициации морфогенеза и дифференцировки зуба. Клеточно-клеточные сигнальные пути и их целевые ядерные факторы были определены в качестве ключевых посредников прогрессивно сложного обмена информацией между эктодермой и эктомезенхимой. Постоянно меняющееся направление обратной сигнализации и реакции клеток между эктодермой и эктомезенхимой позволяет клеткам непрерывно контролировать их относительные пространственные положения и дифференцированные состояния. Наименее понятными из ранних процессов развития зуба являются морфогенез и паттернинг [7]. Из, казалось бы, однородного слоя пероральной эктодермы и подстилающей массы эктомезенхимоцитов, в разных положениях развиваются различные типы и формы зубов. Тип зуба определяется на самом раннем этапе развития, до явного начала морфогенеза. Эти процессы мало изучены и во многом необъяснимы, несмотря на многочисленные исследования ранних клеточных взаимодействий эктодермы-эктомезенхимы и их реакций на позиционные различия в развивающейся челюсти. Многочисленные исследования, выполненные на материале эмбрионов человека, рассматривают развитие зубов, исходя из известных 2-х типов исходных клеток, что, по нашему мнению, является недостаточным.
Цель исследования: изучить развитие и особенности морфогенеза зубов человека на ранних этапах эмбрионального развития.
Материалы и методы исследования
Исследования проведены на эмбрионах и плодах человека (рис. 1) в соответствии с требованиями Минздравмедпрома РФ от 29.04.94 № 82 и согласно номенклатуре клинических лабораторных исследований МЗ РФ (приказ № 64 от 21.02.2000 г.) с учётом положений Хельсинкской декларации (2013). С помощью кластерного анализа исследованы потенциальные однородные подгруппы материала в соответствии с принципами доказательной медицины. Использован классический морфологический метод исследования с окрашиванием срезов гематоксилином и эозином с последующим анализом полученного иллюстративного материала. Ретроспективная оценка результатов проводилась по морфологическим признакам, наблюдавшимся при использовании микроскопа Olympus Bx 52. Исследование проведено с разрешения Этического комитета ФГБОУ ВО ТГМУ и ФГАОУ ВО ДВФУ.


а) б)
Рис. 1. а) Эмбрион человека на стадии 30 сомитов; б) Головной отдел эмбриона человека в период нейруляции. Нативные препараты. Ув. х100
Результаты исследования и их обсуждение
На самом раннем этапе развития человека на стадии 10 и более сомитов (рис. 2) обособляется головной конец зародыша человека и появляется ротовая ямка (рис. 3).
а) б)
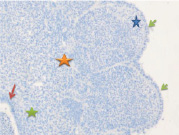
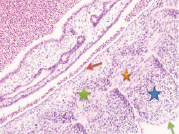
Рис. 2. Эмбрион человека в конце 3-й недели. Стадия а) 10; б) 30 сомитов. 2-х слойная эктодерма туловищного отдела зародыша человека (указана зелеными стрелками), вдается между сомитами более чем на 1/3 зародыша. Голубая, красная и зеленая звёздочки соответственно указывают дерматом, миотом и склеротом). Красные стрелки указывают на энтодерму. а) мезодерма в начале дифференцировки на дерматом, миотом и склеротом; б) выраженная дифференцировка мезодермы на дерматом, миотом и склеротом. Микрофото. Окраска гематоксилином. Увеличение х100
а) б)
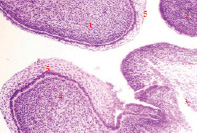
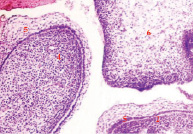
Рис. 3. Полость рта эмбриона человека пяти недель. 1, 2) носовой и медиальный небный отростки формирующейся верхней челюсти; 3) язык; 4) нижняя челюсть, 5) многослойный плоский неороговевающий эпителий. Окраска гематоксилином и эозином. Ув. х200
Наши данные являются убедительным доказательством морфологических отличий эпителия преддверия полости рта и эпителия собственно ротовой полости, имеющих соответственно эктодермальное происхождение, и из выстилки передней кишки.
Первоначально эпителий, выстилающий полость рта, является однослойным, однако уже в начале 5-й недели он превращается в двухслойный, который становится многослойным в конце 5-й, начале 6-й недели (рис. 4).
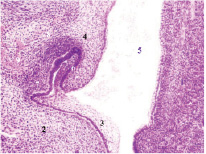
Рис. 4. Полость рта эмбриона человека. Проксимальный отдел пищеварительной трубки: 1) формирующаяся верхняя челюсть; 2) нижняя челюсть, 3) многослойный плоский неороговевающий эпителий; 4) закладка зуба, 5) проксимальный отдел пищеварительной трубки; 6) эмалевый орган; 7) зубной сосочек; 8) зубной мешочек; 9) формирующаяся кость нижней челюсти. Окраска гематоксилином и эозином. Ув. х200
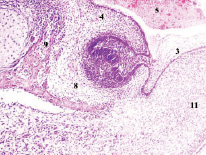
Рис. 5. Полость рта эмбриона человека: 1) формирующаяся нижняя челюсть, 2) цилиндрический эпителий; 3) многослойный плоский неороговевающий эпителий; 4) закладка зуба, 5) проксимальный отдел пищеварительной трубки; 6) эмалевый орган; 7) зубной сосочек; 8) зубной мешочек; 9) формирующаяся кость зубной альвеолы; 10) Меккелев хрящ; 11) губа. Окраска гематоксилином и эозином. Ув. х200
Наши результаты показали, что формирующийся эмалевый орган имеет несимметричное строение вследствие разных топографических особенностей и взаимоотношений его пролиферирующих структур с эктомезенхимой, прилежащей к эктодермальному эпителию, и части эмалевого органа, располагающейся вблизи выстилки передней кишки. Наклонный вектор роста эмалевого органа также связан с асимметричной пролиферацией клеток. Идентифицируются участки образующейся костной ткани нижней челюсти. Базальный слой эктодермального эпителия, врастающего в мезенхиму нижней челюсти, образует внутренний и наружный слои эмалевого колпачка. В этот период отмечается неодинаковая толщина слоев клеток эмалевого органа, наибольшие значения которой отмечаются со стороны полости рта. Зубной мешочек в меньшей степени окружает формирующиеся структуры эмалевого органа со стороны преддверия полости рта. Это может быть связано с тем, что формирование эмали зуба с внутренней стороны полости рта и со стороны преддверия полости рта имеет отличия в развитии, как со стороны клеточной индукции, так и последующей дифференцировки клеток. D. Kaliboviс Govorko, T. Beсiс, (2010) с соавторами показали, что экспрессия Ki-67-маркера пролиферации, Bcl-2 и Bax протеина имеет значение в развитии зуба [8]. Асимметрия сохраняется и в последующую неделю (рис. 5).
Зубной стебелёк, или шейка зуба, покрыт со стороны преддверия рта базальным слоем эктодермальных кератиноцитов, со стороны собственно полости рта базальные клетки граничат с цилиндрическим эпителием проксимального отдела пищеварительной трубки. Граница перехода чётко идентифицируется, возможно, это связано с ингибированием пролиферации за счёт сигнальных молекул, секретируемых цилиндрическим эпителием и внутренними клетками зубного мешочка, распространяющимися на 5-й неделе до границы наружных эмалевых клеток и цилиндрического эпителия прoксимального отдела пищеварительной трубки
Известно, что головной конец зародыша не содержит мезодермы, поэтому мезенхима в головном отделе представлена двумя типами: эктомезенхимальной и мигрирующей из нервного гребня [9]. Таким образом, в морфогенезе зуба человека участвует большее количество клеток, чем принято считать на современном этапе.
Нами отмечено, что эмалевый орган содержит многочисленные хромофобные клетки, имеющие вытянутую веретеновидную форму. Их отростки пронизывают слой внутренних клеток эмалевого органа и формируют мембрану, обособляющие эмалевый орган от зубного сосочка, вдающегося в него. Физиологическое значение мембраны может быть связано с ограничением миграции клеток энтомезенхимы. Вторая мембрана идентифицируется на границе слоя энамелобластов и пульпы эмалевого органа. Наиболее крупные хромофобные клетки располагаются в той части формирующегося зуба, которая является наружной, обращённой в преддверие рта.
Хромофобные клетки образуют капсулу вокруг зубного сосочка, располагаются в его паренхиме и представляют большую часть клеток, формирующих зубной мешочек. Нами отмечено присутствие крупных хромофобных клеток на границе места перехода многослойного плоского эпителия и его базальных кератиноцитов в цилиндрический эпителий проксимального отдела развивающейся пищеварительной трубки. В эпителии слизистой оболочки разных отделов полости рта, который в дальнейшем превратится в частично ороговевающий и неороговевающий, возникают различия в экспрессии цитокератинов. Возможно, это связано с тем, что в неороговевающем эпителии, выстилающем ротовую полость, базальные кератиноциты располагаются на мембране, представленной хромофобными веретеновидными клетками, происхождение которых может быть связано с нервным гребнем, или эктомезенхимой. Следует отметить, что мембрана из веретеновидных клеток лучше идентифицируется в эпителии и вокруг эмалевого органа, отсутствует в проксимальном отделе пищеварительного канала. S. Panneer Selvam, I. Ponniah (2018) пришли к выводу, что экспрессия амелобластина в зародышах зуба человека связана с дифференцировкой и минерализацией [10].
Несмотря на известные модели экспрессии в геноме высоко пролиферативных дифференцирующихся амелобластов и одонтобластов на ранних и поздних стадиях развития эмалевого органа генов IGF-2, IGF-1R, IGF-2R и PTEN, имеющих важное значение в морфогенезе коронки зубов человека, выращивание искусственных зубов in vitro пока затруднено и не представляется возможным не только для человека, но и для животных [11, 12]. Неэффективное использование известных сигнальных молекул, ростовых факторов и биологически активных веществ в клеточных технологиях по выращиванию зубов диктуют изучение развития зубов и поиск новых не известных на современном этапе данных.
Заключение
Таким образом, развитие зуба является результатом последовательных и взаимных взаимодействий между эпителием полости рта и нейрональной мезенхимы. В нашем исследовании основное внимание мы уделили неамелобластным слоям ЭО: поверхностным слоям, звездчатому ретикулуму пульпы эмалевого органа и наружному эмалевому эпителию, отметив, что в структуру эмалевого органа произошла миграция эктомезенхимоцитов веретеновидной формы, хромофобных, простирающихся перпендикулярно мембране энамелобластов. Открытие новых фактов в формировании корня зуба, коронкового дентина и эмали указывает на возможность утверждения новой концепции: коронка зуба и корень имеют различные механизмы индукции направления дифференцировки и специализации клеток. Эти данные показывают, что слои неамелобластов ЭО играют несколько ролей во время одонтогенеза, включая поддержание нескольких резервуаров стволовых клеток, играют важную роль во время морфогенеза корня зуба, стабилизирующую функцию для слоя амелобластов. Формирование зубных структур или зубов, как органов, в эксперименте in vitro зависит от знания стволовых клеток и требует взаимодействия всех межклеточных и молекулярных факторов, которые приводят к образованию не только специфичных для зубов твердых тканей, дентина, цемента и эмали, но и пульпы. Хотя мезенхимальные стволовые клетки различного происхождения были широко изучены в их способности образовывать дентин in vitro, информации об успешном использовании эпителиальных стволовых клеток в выращивании зубов пока нет. Одонтогенный потенциал находится в зависимости от эпителиальных стволовых клеток, необходимых как для инициации образования зуба, так и для производства эмалевого матрикса. Эмбриональные постнатальные или даже взрослые стволовые клетки обладают огромным регенеративным потенциалом, но их применение в стоматологической практике все еще проблематично и ограничено из-за различных неизвестных параметров развития зубов. Недостаточность информации о клеточных взаимодействиях в развитии зубов человека влияет на высокий риск отторжения и непредсказуемость поведения стволовых клеток, длительный период прорезывания зубов, не обеспечивают морфогенез заданной формы и соответствующей структуры коронки.
В современных работах о развитии зубов у человека в реальном развитии и in vitro показано, что постнатальные стволовые клетки пульпы человека, как и стволовые клетки эпителия слизистой рта, не обладают одонтогенным потенциалом или одонтогенной компетентностью. Мы связываем это с отсутствием в этих процессах хромофобных веретеновидных клеток, присутствие которых необходимо для индуцированного выращивания зубов.
Выводы
Результаты, полученные на материале эмбрионов человека, демонстрируют возможность сохранения одонтогенного потенциала в зубных эмбриональных тканях человека с определённым ансамблем клеток и будут иметь значение в перспективе в биоинженерных технологиях выращивания зубов человека. Технологии тканевой инженерии и регенеративной медицины, как перспективные методы лечения в стоматологии, обязательно должны учитывать хромофобные веретеновидные клетки, мигранты из нервного гребня и эктомезенхимы, участвующие в развитии зубов человека на самых ранних этапах эмбрионального развития. Возможно, эти клетки являются главными координаторами структуризации, дифференцировки и специализации формирующихся зачатков зубов у эмбрионов человека.
Работа выполнена при поддержке Научного фонда ДВФУ, в рамках государственного задания 17.5740/2017/6.7.
Библиографическая ссылка
Рева И.В., Ямамото Т., Восканян О.Г., Барановская И.А., Одинцова И.А., Верин В.К., Кожухарь В.Г., Ким А.Р., Рева Г.В. ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ ЗУБОВ ЧЕЛОВЕКА // Международный журнал прикладных и фундаментальных исследований. 2019. № 3. С. 44-49;URL: https://applied-research.ru/ru/article/view?id=12681 (дата обращения: 04.07.2026).
DOI: https://doi.org/10.17513/mjpfi.12681