

В связи с повышением общественного интереса к творческой деятельности человека возросло количество научных исследований, посвященных расширению сферы восприятия, в частности развитию доминирующего типа восприятия и синестезий [1–3].
Целью данной работы являлись поиск и определение алгоритма деятельности мозга в процессе проявления отдельных преобладающих видов восприятия: зрительного, слухового, соматосенсорного у лиц с доминирующей сенсорной модальностью – визуалов, аудиалов, кинестетиков. Разработка нейрофизиологического инструмента на основе альфа-стробирования сенсорных сигналов [4, 5] дает возможность исследовать относительно устойчивые сенсорные системы саморегуляции с участием эмоциональных образований, имеющих связь с памятью. При этом стробирование можно рассматривать как один из механизмов в развитии системы саморегуляции, так как оно позволяет осуществлять комбинирование восприятия и действий в создаваемых им интервалах времени. Использование такого инструмента позволяет достаточно детально изучать условия развития доминирующего типа восприятия и участие в этом процессе синестезий.
В настоящем исследовании синестезии рассматриваются как способность преобразования сигналов, поступающих в мозг по сенсорным каналам одной модальности, в образы другой сенсорной модальности. Подобные синестезии могут вносить качественные изменения в систему восприятия, поскольку могут переводить слабые неосознаваемые ощущения одних органов чувств в осознаваемые другой, более развитой у конкретного человека, системой восприятия. Данный тип синестезии характерен для экстрасенсорного восприятия и может возникать на фоне особых эмоциональных состояний, например таких, как эмпатия (сопереживание) [6].
Для выполнения задач, направленных на изучение синестезий и доминирующих типов восприятия, необходимо ответить на следующие вопросы, имеющие непосредственное отношение к структуре исследования.
1. Каковы особенности спектральных характеристик альфа-ритма и их распределения на поверхности коры у визуалов, акустиков, кинестетиков? Как у них локализуются источники происхождения альфа-ритма?
2. Как глубинные источники происхождения альфа-ритма проецируют активность в кору головного мозга?
3. Является ли постоянно доминирующая в коре мозга зона первичного восприятия у визуала, аудиала или кинестетика проявлением синестезий по отношению к остальным зонам сенсорного восприятия?
Материалы и методы исследования
Альфа-стробирование сенсорных сигналов мозга при доминирующем типе восприятия исследовано в условиях, оптимальных для проявления альфа-активности у испытуемых: в состоянии спокойного бодрствования, психической и мышечной релаксации с закрытыми глазами. Эти условия позволили рассматривать индивидуальные показатели альфа-ритма как наиболее адекватные и присущие каждому участнику исследования для отражения готовности к активной мозговой деятельности и восприятию сенсорных сигналов. Не исключено, что при этом стабильность рассматриваемых показателей обеспечивается процессами не только в сенсорных центрах первичного восприятия в коре, но и в структурах контроля и регуляции эмоций и памяти в лимбической системе, а также в системе восходящей неспецифической активации в структурах ствола мозга и таламуса.
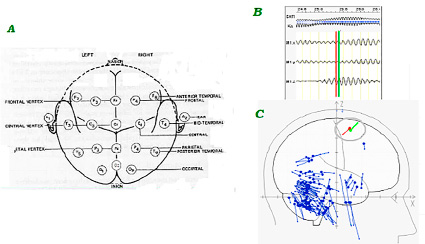
Рис. 1. A. Стандартное расположение электродов на поверхности головы по схеме 10/20. B. Регистрация альфа-ритма в проекциях головы XYZ (соответственно M1x, M1y, M1z) синхронно с вычислениями эквивалентных дипольных источников и их векторов. Данные, относящиеся к мозжечку, в статье не рассматриваются. С. Два эквивалентных дипольных источника (в овале) с противоположно направленными векторами: красный в сторону положительной поляризации диполя в области гиперполяризации нейронов, зеленый – в сторону отрицательной поляризации в области деполяризации нейронов. Моменты вычисления этих источников совпадают с красной и зеленой линиями на рис. В, пересекающими линии альфа-ритма с интервалом, равным полупериоду альфа-волны
В исследовании участвовала группа испытуемых, занимающихся интеллектуальным трудом (16 женщин и 14 мужчин), в возрасте от 30 до 60 лет. Для отбора лиц с доминирующим типом восприятия (визуалов, аудиалов, кинестетиков) была проведена регистрация ЭЭГ у 120 чел. в состоянии спокойного бодрствования, психической и мышечной релаксации, с закрытыми глазами. Отбор испытуемых проводился на основе показателей спектральных характеристик альфа-ритма в первичных зонах коры основных сенсорных модальностей: зрительной, слуховой, соматосенсорной [5]. При наличии максимумов спектральной мощности альфа-ритма частотой от 7 Гц до 13 Гц с шагом 1 Гц в одной из первичных сенсорных зон коры делали вывод о доминировании зрительного восприятия (поля по Бродману 17, 18, 19) у визуалов, слухового (поля по Бродману 41, 42, 20, 21, 22, 37) – у аудиалов, сенсомоторного (поля по Бродману 2, 3, 4, 6) – у кинестетиков. В группу «Баланс» были включены испытуемые, у которых отсутствовало доминирование одной из сенсорных модальностей. Основные типы восприятия проявлялись равномерно, и это отражалось в повышении спектральной мощности альфа-ритма в первичных центрах сенсорного восприятия на свойственной ему частоте согласно альфа-стробированию: 7–9 Гц у кинестетиков и аудиалов, 9–11 Гц у визуалов, 11–13 Гц у аудиалов.
Регистрацию ЭЭГ проводили на 24-канальном нейровизоре NVX24 производства ООО «Медицинские Компьютерные Системы» с использованием стандартных монополярных отведений в соответствии с принятой международной схемой 10–20 (рис. 1, А) [7].
Исследования проводились в стандартных, одинаковых для всех испытуемых условиях ограничения электромагнитных наводок, слуховых и других сенсорных помех.
Для обработки и анализа данных регистрации электрической активности мозга – электроэнцефалограммы (ЭЭГ) использованы пакеты компьютерных программ: топографического картирования электрической активности мозга Brainsys Neuro-KM [8] и Программа трехмерной локализации источников электрической активности головного мозга BrainLoc [8, 9].
Применены методики нейрокартирования в виде распределения показателей мощности частотного спектра альфа-ритма по отдельным областям на поверхности полушарий мозга. Использован статистический анализ для оценки достоверности получаемых данных.
С целью выявления функциональной архитектуры доминирующего сенсорного восприятия вычисляли распределение эквивалентных токовых диполей (ЭТД) генераторов альфа-ритма частотой 7–13 Гц с шагом 1 Гц в коре и в глубинных структурах мозга (рис. 1, C). Локализацию ЭТД определяли в каждый момент времени, равный дискретному интервалу отсчета данных при вводе в компьютер. Вопрос о соответствии локализации ЭТД в глубинных структурах мозга той биоэлектрической активности, которая регистрировалась на поверхности головы, решался специальным алгоритмом, в котором рассматривались электрические свойства мозга как объемного проводника [9, 10]. Для анализа применяли однодипольную модель ЭТД. Одновременно программы вычисляли параметры анатомической локализации ЭТД, включая корковые поля Бродмана и отдельные образования серого и белого вещества в подкорковых областях [11].
При интерпретации результатов работы программы учитывали [9, 10], что вычисляемый для анализируемых одномоментных сечений ЭЭГ токовый диполь является не реальным источником потенциалов ЭЭГ, а эквивалентным, результирующим источником электрически активной в данный момент времени области мозга. ЭТД характеризует суммарную электрическую активность нервных клеток, вовлеченных в текущий момент времени в процесс электрического возбуждения, и расположен в электрическом центре этой области. Поэтому коэффициент дипольности, как показатель того, насколько точно вычисленная эквивалентная дипольная модель описывает пространственное распределение потенциалов на поверхности головы, был использован на уровне высоких значений КД => 0, 97. При этом ЭТД источников альфа-ритма с высоким коэффициентом дипольности (КД => 0, 97) могут рассматриваться как указатели на наиболее активные локальные зоны, запускающие последовательное перемещение волновых процессов альфа-колебаний. Высокий уровень дипольности ЭТД показывает, что интенсивность электрического поля генераторов альфа-ритма достаточна, чтобы достичь поверхности коры.
Результаты исследования и их обсуждение
Так как в норме повышение амплитуды и спектральной мощности альфа-ритма отражает готовность к функциональной активности, то доминирующий тип восприятия определяли именно в состоянии покоя и готовности путем выявления максимумов спектральной мощности альфа-ритма в первичных сенсорных зонах коры головного мозга. При этом учитывалось, что такая локализация выделенных макроскопических сетей в состоянии покоя совпадает или преобразуется в относительно устойчивую, но сходную с покоем, модификацию во время активной деятельности, например при наблюдении за внешними событиями или при их воспоминаниях [12, 13].
Усредненные распределения спектральной мощности альфа-ритма в коре мозга, полученные по отдельным группам визуалов, аудиалов, кинестетиков, представлены на рис. 2. Эти распределения в каждой группе имеют свои особенности, но общим является то, что у лиц с доминирующим типом восприятия альфа-ритм имеет максимум спектральной мощности в первичной зоне коры доминирующей модальности сигналов на всех частотах от 7 до 13Гц. Особенно четко эта особенность проявляется при сопоставлении с распределением спектральной мощности альфа-ритма в коре головного мозга у лиц с балансированным восприятием сенсорных сигналов всех модальностей (рис. 2, БАЛАНС).
Согласно представлениям об альфа-стробировании [5] ритмы частотой 9–12 Гц имеют отношение к зрительному восприятию, и максимальная спектральная мощность этих ритмов концентрируется в зрительных отделах коры затылочных областей (рис. 2, БАЛАНС). Но у визуалов максимальная спектральная мощность альфа-ритма наблюдается в зрительной зоне затылочной коры в более широком диапазоне частот от 7 до 13 Гц (рис. 2, ВИЗУАЛ). Этот признак говорит о повышенной готовности зрительных отделов коры к восприятию сигналов разных модальностей в рамках альфа-стробирования и является отличительной особенностью активности мозга у визуалов.
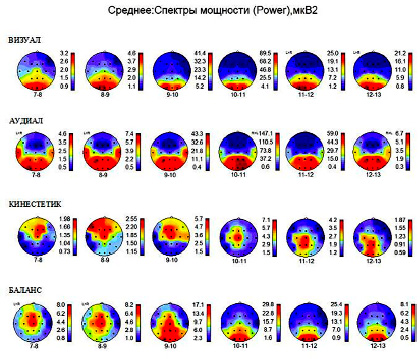
Рис. 2. Карты распределений спектральной мощности альфа-ритма, усредненной по отдельным группам испытуемых с доминирующим типом восприятия: визуалов (ВИЗУАЛ), аудиалов (АУДИАЛ), кинестетиков (КИНЕСТЕТИК) и с отсутствием доминирования (БАЛАНС). В нижней части каждой карты указана частота альфа-ритма, справа цветовая шкала уровня спектральной мощности мкВ2. Зоны отведений ЭЭГ см. на рис. 1, А
В группе аудиалов (рис. 2, АУДИАЛ) максимумы мощности альфа-ритма наблюдаются в височных областях, чаще в сочетании со зрительными зонами. В группу вошли участники, ежедневно творчески занимающиеся прослушиванием музыкальных произведений или созданием новых мелодий.
Можно предположить, что постоянная творческая деятельность стимулирует именно те системы мозга, которые обеспечивают наиболее широкое восприятие специфических сигналов творчества. К ним относятся восходящие системы неспецифической и эмоциональной активации мозга, многочисленные связи и взаимодействия активных центров, в состоянии готовности которых наиболее выражена мощность альфа-ритма.
Особенностью данной группы является повышенная мощность альфа-ритма на всех рассматриваемых частотах не только в центрах слухового восприятия в височной области, но также и в центрах зрительного восприятия в затылочной области и в теменных ассоциативных зонах. Могут ярко проявляться аудио-визуальные синестезии.
В группе кинестетиков (рис. 2, КИНЕСТЕТИК) в распределениях средней мощности альфа-ритма максимум наблюдается в сенсомоторной области на всех частотах в диапазоне от 7 до 13 Гц. Некоторое смещение максимума в ассоциативную зону теменной области наблюдается только на частоте 11–13 Гц на фоне общего снижения показателей спектральной мощности на поверхности полушарий.
При отсутствии доминирующего типа восприятия (рис. 2, БАЛАНС) активность альфа-ритма в первичных центрах основных сенсорных модальностей (зрительной, слуховой, кинестетической) распределяется соответственно частотному альфа-стробированию.
Возникает вопрос, каким образом формируются каналы передачи сенсорной информации как доминантной, так и недоминантной модальности у лиц с доминирующим типом восприятия?
Особенности альфа-стробирования сенсорных сигналов при доминирующем типе восприятия рассмотрим на отдельных примерах.
Визуалы воспринимают мир посредством зрительных образов [3]. Доминирующий орган чувств у них – глаза. Другие формы оценки мира у таких людей тоже развиты, но им проще и удобнее обрабатывать поступающую информацию именно через зрение.
Особенности нейронной активности мозга у визуала показаны на примере испытуемого З. (рис. 3). Представленные карты поверхности мозга с распределениями спектральной мощности альфа-ритма с шагом 1 Гц показывают, что максимумы распределений наблюдаются в зрительных зонах затылочной коры обоих полушарий на всех частотах альфа-колебаний от 7 до 13 Гц (рис. 3, A, B). При этом в зрительных зонах коры, преимущественно в полях 17, 18, 19 и 7 по Бродману, наблюдается повышенная концентрация ЭТД генераторов альфа-колебаний 7–13 Гц с высоким коэффициентом (КД => 0, 98) дипольности (рис. 3, C). Особенно плотная локализация ЭТД в этих зонах наблюдается на частоте альфа-ритма 9–10 Гц и 10–11 Гц. У визуалов распределения ЭТД в структурах лимбической области и восходящей неспецифической активации зависят от частоты альфа-стробирования. В данном примере на рис. 3, Е, наблюдается объединение структур восходящей неспецифической активации (Varolius), лимбической системы (поля Бродмана 23, 30, 31) и зрительной зоны (поля Бродмана 17, 18, 19) в систему каналов на частоте 7–8 Гц.
В объединение на частоте 11–12 Гц добавляются поля Бродмана 7, 29 и 37. На частоте 12–13 Гц дополнены влияния системы восходящей неспецифической активации мозга (Mesencephalon, Thalamus, Substantia Nigra, Nucleus Ruber). Таким образом, у визуалов сохраняется система передачи сигналов по каналам соматосенсорной модальности (7–8 Гц) или слуховой (11–13 Гц и частично 7–8 Гц), которые отличаются от других не только частотной специфичностью, но и анатомическими структурами.
Кроме того, у визуалов вектора ЭТД генераторов альфа-волн в каналах передачи сенсорных сигналов любой, не только зрительной, модальности направлены преимущественно к доминирующему центру зрительного восприятия (рис. 3, D). И в этом заключается важное отличие доминирующего восприятия от балансированного, при котором векторы ЭТД источников альфа-ритма определенной модальности имеют направления преимущественно в сторону центра восприятия той же сенсорной модальности [5].
Отсюда следует (рис. 3, A, B, C, D), что обусловленная альфа-стробированием частотная специфичность каналов передачи сенсорных сигналов каждой модальности создает возможность взаимодействия сенсорных сигналов зрительного спектра с соматосенсорными и слуховыми сигналами, исходящими из систем соответствующего сенсорного восприятия.
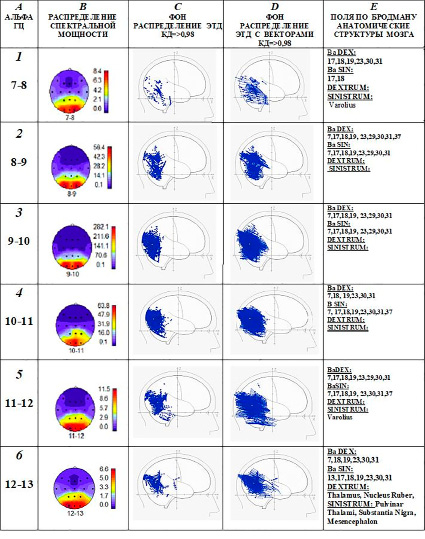
Рис. 3. Пример: испытуемый З. – визуал. А, В (1, 2, 3, 4, 5, 6) – максимумы спектральной мощности альфа-ритма в зрительных зонах затылочной области коры. C, Е (1, 2, 3, 4, 5, 6) – преобладание распределений ЭТД в зрительных зонах и в поясной извилине; 1, 5, 6 – ЭТД наблюдаются также в структурах системы неспецифической восходящей активации. D (1, 2, 3, 4, 5, 6) – ЭТД с векторами, направленными в сторону затылочной области. Обозначения столбцов: A – частота альфа-ритма в Гц; B – схемы коры мозга (зоны отведений ЭЭГ см. на рис. 1) с распределением спектральной мощности альфа-ритма, справа цветовая шкала в мкВ2, под схемой указана частота альфа-ритма; C – локализация ЭТД генераторов альфа-ритма в структурах мозга в сагиттальной проекции; D – то же, что и в п. C, с добавлением изображений векторов ЭТД; E – локализация ЭТД генераторов альфа-ритма в анатомических структурах мозга, BaDEX, BaSIN – поля Бродмана в правом и левом полушариях, DEXTRUM, SINISTRUM – глубинные структуры правого и левого полушарий. 1, 2, 3, 4, 5, 6 – обозначения частот альфа-ритма для изображений по горизонтали
При этом функция стробирования проявляется в процессе сканирования электрическими альфа-волнами, которые могут распространяться непосредственно в средах мозга, влияя на состояние возбудимости мембран отдельных элементов нейронных сетей [10, 13–15]. В рассматриваемом случае скопление ЭТД источников альфа-ритма частотой 7–8 Гц наблюдается в поясной извилине. При этом проявляется однонаправленность векторов ЭТД в сторону зрительной коры мозга. Таким образом, благодаря альфа-стробированию сигналы соматосенсорной модальности в поясной извилине могут проецироваться альфа-волной в зрительную зону коры. Так как зрительная кора способна работать только со зрительными образами, то соматосенсорная информация преобразуется в зрительную. В этом случае можно говорить о синестезии, то есть о преобразовании сигналов восприятия одной модальности ощущений в образы другой модальности.
Если одновременно альфа-ритм более низкой частоты 7–9 Гц и более высокой 11–13 Гц также имеет максимум мощности в затылочной области коры, то это означает возможность возникновения кинестетико-визуальных или аудио-визуальных синестезий. У визуала в случае более слабых почти неосознаваемых слуховых (альфа-стробирование 12–13 Гц) или кинестетических (альфа-стробирование 7–9 Гц) сигналов в центрах зрительной коры благодаря повышенной возбудимости возможно преобразование в более яркие и четкие зрительные образы. В определенных условиях, когда передача по нейронным путям сигналов от органа чувств будет иметь подпороговое значение и не осознаваться, можно допустить, что анализатор более развитой модальности ощущений с помощью эфаптической связи отсканирует информацию и создаст осознаваемый образ.
Для аудиалов характерны следующие признаки: хороший слух, любовь к музыке, высокая требовательность как к своей, так и к чужой речи. Аудиалы больше других ориентированы на общение, им необходима полная тишина при сосредоточении на чем-либо, они имеют хорошую слуховую память [3].
У аудиала А. (рис. 4, А, В) спектральная мощность альфа-ритма на всех частотах увеличена в височных областях в первичных и вторичных зонах слухового восприятия, включая зоны Вернике, а также в ассоциативных теменных областях и в зонах зрительного восприятия затылочной области. Такое распределение повышенной мощности альфа-ритма в коре в состоянии покоя, по-видимому, отражает готовность к наиболее часто выполняемым видам деятельности нейросетей. Как показано [12, 13], индивидуальные нейросети относительно стабильны по структуре, хотя имеют специфическую динамику активной деятельности, а при сопоставлении с сетями состояния покоя совпадают по локализации. Известно, что полимодальное восприятие и многочисленные ассоциации между слуховыми, зрительными и соматосенсорными образами характерны для музыкального творчества в процессе создания новых или исполнения готовых музыкальных произведений. В эту группу были включены представители творческой музыкальной профессии.
ЭТД с высоким уровнем дипольности (КД => 0, 98) локализуются в слуховых зонах височной области, вторичных и третичных, а также в ассоциативных зонах зрительного и соматосенсорного восприятия коры больших полушарий, в поясной извилине и в глубинных структурах – гиппокампе, хвостатом ядре, таламусе, среднем мозге (рис. 4, Е). Необходимо отметить, что ЭТД генераторов альфа-ритма частотой 7–13 Гц с шагом 1 Гц распределяются в слуховых и в зрительных зонах коры, в поясной извилине, в Mesencephalon. Это структуры, которые определяют доминирование слухового и зрительного восприятия в данной творческой группе. Однако в отдельных глубинных образованиях мозга выделяются ЭТД источников альфа-волн в более узких диапазонах частот, соответствующих разным модальностям. Векторы большинства ЭТД направлены в стороны височных и затылочных областей. Они указывают на возможность переноса активности эфаптическим путем. Появляются условия возникновения синестезии – преобразования сигналов одной модальности в чувственный образ другой модальности, например, слухового образа на основе зрительных или соматосенсорных сигналов. В более расширенном представлении синестезия – это явление восприятия, при котором стимуляция одного сенсорного или когнитивного пути приводит к непроизвольным переживаниям во втором сенсорном или когнитивном пути [1, 2].
Кинестетик овладевает информацией, полагаясь на свои физические ощущения, на органы осязания. Кожные, мышечные ощущения, состояние внутренних органов и собственная реакция на них имеют первостепенное значение для человека с превалирующим кинестетическим каналом. Такие люди очень тонко и подлинно распознают чувства и эмоции [3].
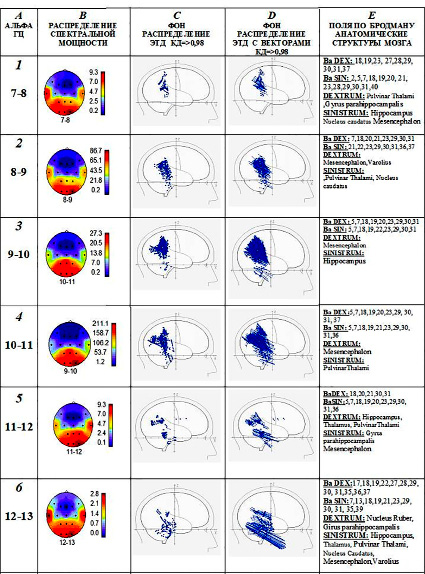
Рис. 4. Пример: испытуемый К. – аудиал. А, В (1, 2, 3, 4, 5, 6) – повышение спектральной мощности альфа-ритма наблюдается в слуховых зонах височной области, в зрительных зонах затылочной области и в ассоциативных зонах теменной области. C, Е (1, 2, 3, 4, 5, 6) – ЭТД альфа-ритма распределяются в слуховых, в зрительных зонах, в поясной извилине, в Mesencephalon, а также 1, 2, 4, 5, 6 – Pulvinar Thalami; 1, 3, 5, 6 – Hippocampus; 1, 5, 6 – Girus Parahippocampalis; 1, 2, 6 – Nucleus Caudatus; 2, 6 – Varolius. D (1, 2, 3, 4, 5, 6) – ЭТД с векторами, направленными в сторону слуховых зон височной области. Обозначения A, B, C, D, E и 1, 2, 3, 4, 5, 6 как и на рис. 3
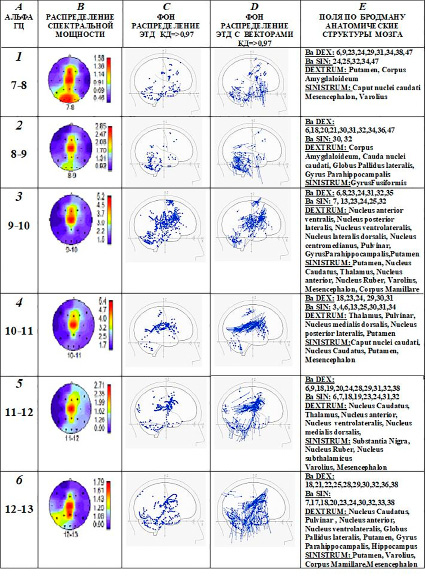
Рис. 5. Пример: испытуемый Б. – кинестетик. А, В (1, 2, 3, 4, 5, 6) – максимум спектральной мощности альфа-ритма наблюдается в центральной сенсомоторной зоне. C, Е (1, 2, 3, 4, 5, 6) – ЭТД альфа-ритма распределяются в премоторной зоне коры, в поясной извилине, Nucleus Caudatus, а также 1, 3, 4, 6 – Putamen; 1, 2 – Corpus Amygdaloideum; 2, 6 – Globus Pallidus; 2, 3, 6 – Girus Parahippocampalis; 3, 4, 5, 6 – Thalamus nuclei; 1, 3, 4, 5, 6 – Mesencephalon; 1, 3, 5, 6 – Varolius. D (1, 2, 3, 4, 5, 6) – имеются ЭТД с векторами, направленными в сторону сенсомоторной области. Обозначения A, B, C, D, E и 1, 2, 3, 4, 5, 6 как и на рис. 3.
У кинестетика спектральная мощность альфа-ритма всего диапазона частот от 7 Гц до 13 Гц имеет максимум в доминирующей сенсомоторной области коры головного мозга (рис. 5, А, В). В этой области отдельные наиболее активные зоны представлены ЭТД источников альфа-волн с высоким коэффициентом дипольности от 0,97 и выше. ЭТД локализуются в первичной зоне соматосенсорного восприятия 3 и в моторной зоне 4 по Бродману, а также в ассоциативных и вторичных моторных или двигательных зонах коры – в полях Бродмана 6, 7, 8, 9, 13 (рис. 5, C, D, E). Активные ЭТД с КД => 0, 97 концентрируются также в отдельных зонах поясной извилины, где представлены поля 23, 24, 29, 30, 38 по Бродману, которые связаны с развитием эмоций и исполнением инстинктивного поведения [11]. ЭТД с высокими значениями дипольности одновременно распределяются в отдельных ядрах таламуса, миндалевидного комплекса, хвостатого ядра, скорлупы, бледного шара, гиппокампа, парагиппокампальной извилины (рис. 5, C, D, E). Вектора эквивалентных токовых диполей генераторов альфа-ритма в этих перечисленных зонах направлены преимущественно в сторону центральных (сенсомоторных) и передних областей мозга. Такая направленность векторов ЭТД создает дополнительные возможности сканирования и переноса сигналов эфаптическим путем из каналов одной сенсорной модальности, включая лимбическую и неспецифическую восходящей активации системы, в первичный центр другой чувствительности. В таком случае телесные ощущения кинестетика могут соответствовать зрительным образам или слуховым сигналам. В результате соматосенсорные или телесные ощущения могут быть более контрастными и осознаваемыми, чем слуховые или зрительные образы.
Кроме того, в создании сложных образов на основе зрительной и соматосенсорной информации большое значение имеет поле Бродмана 7. Оно является третичным проекционным полем, ассоциативной зоной в месте перекрытия различных анализаторов. Здесь происходит интеграция различных сигналов, формирование соответствующих ощущений, осуществление сложных аналитико-синтетических функций.
Подобно тому как при активации первичного зрительного центра возбуждаются генераторы ритмического волнового процесса, сканирующего сенсорную кору и распространяющегося с периодом альфа-ритма [13–15], в настоящей работе сканирующие процессы отражаются в интервалах альфа-стробирования, которые определяют частотную специфичность каждой сенсорной модальности не только в первичных сенсорных зонах, но и в структурах лимбической и неспецифической восходящей систем.
В рассматриваемой нами функциональной архитектуре альфа-стробирования каждая альфа-волна переносит эфаптическим способом состояние отдельных структур лимбической системы в ту область, куда направлен вектор отрицательного знака (рис. 1, В, С). У лиц с доминирующим типом восприятия наблюдается преимущественное направление отрицательных векторов в первичную зону доминантной модальности независимо от того, к системе какой модальности принадлежит альфа-волна. При этом деполяризующий (возбуждающий) фронт альфа-волны является активным передатчиком результатов сканирования, а векторы, направленные к положительному полюсу ЭТД, показывают движение альфа-волны гиперполяризующим (тормозным) фронтом воздействия на структуры мозга, находящиеся вне доминирующей зоны восприятия, тем самым создавая контрастные условия для концентрации доминирующей активности.
Выводы
1. У лиц с доминирующим типом восприятия сохраняется частотная специфичность по отношению к сенсорным сигналам каждой отдельной модальности: зрительной 9–11 Гц, слуховой 12–13 Гц, соматосенсорной 7–9 Гц, что подтверждает наличие процесса альфа-стробирования. Однако именно у доминантов основной особенностью распределения спектральной мощности альфа-ритма в коре мозга является локализация максимумов в первичной зоне доминирующей сенсорной модальности сразу во всем диапазоне частот от 7 Гц до 13 Гц.
2. При доминирующем типе восприятия эквивалентные токовые диполи генераторов альфа-ритма с высоким коэффициентом дипольности КД => 0,97 локализуются в коре головного мозга преимущественно в первичной зоне доминирующей сенсорной модальности. Повышенная концентрация ЭТД генераторов альфа-волн наблюдается также в тех проводящих системах и структурах лимбической системы, отвечающих за контроль и регуляцию эмоций и памяти, и системы восходящей неспецифической активации мозга, которые имеют отношение к доминирующему восприятию.
3. Благодаря альфа-стробированию в структурах лимбической системы возникают эквивалентные токовые диполи источников альфа-ритма в частотных диапазонах соответствующих модальностям – зрительной, слуховой, соматосенсорной. Векторная направленность этих диполей в сторону доминирующей первичной зоны в коре указывает на возможность эфаптической передачи альфа-волной сигналов возбуждения от образований лимбической системы, которые участвуют в происхождении эмоциональных реакций и запоминании зрительных, слуховых, соматосенсорных сигналов и образов.
4. Предполагается, что в центре доминирующего восприятия происходит возбуждение альфа-волной, несущей сигналы от структур лимбической системы, и формируется образ доминирующей сенсорной модальности. Таким путем происходит преобразование сигналов любой сенсорной модальности в сигналы и образы доминирующей модальности. Способность преобразования сигналов, поступающих в мозг по сенсорным каналам одной модальности, в образы другой сенсорной модальности рассматривается как синестезия.
Библиографическая ссылка
Кузяев А.Е., Коёкина О.И. АЛЬФА-СТРОБИРОВАНИЕ СЕНСОРНЫХ СИГНАЛОВ МОЗГА И СИНЕСТЕЗИИ ПРИ ДОМИНИРУЮЩЕМ ТИПЕ ВОСПРИЯТИЯ // Международный журнал прикладных и фундаментальных исследований. 2022. № 11. С. 19-29;URL: https://applied-research.ru/ru/article/view?id=13462 (дата обращения: 12.06.2026).
DOI: https://doi.org/10.17513/mjpfi.13462