

Н.И. Вавилов [4] выявил, что «Генетически близкие виды и роды характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм у других видов и родов». Урманцев Ю.А. [35] утверждал, что «Любой мыслимый объект – система, и любой объект непременно должен принадлежать хотя бы одной системе объектов того же рода». И это «позволяет сделать ряд новых предсказаний и обобщений, открыть новые факты, законы и явления, найти оригинальные связи и решения, обнаружить и исправить ошибки прежних исследований». Априори любой объект мироздания – система таксон, в таксоне таксонов и хотя бы по нескольким признакам гомологичен, аналогичен или идентичен любому другому объекту мироздания – таксону. Это положение и позволяет объединять любые объекты в таксоны разного ранга системности и познавать все объекты мироздания.
Эволюция материи, как объект-система S объединяет ряд таксонов: эволюция частиц, атомов, молекул, организмов и т.д. Каждая из этих объект-систем должна обладать, частью или всеми системно-структурными признаками, подчиняться всем или части системных отношений и законам, реализованным в системе объектов рода S.
Рассмотрим малую периодичность – «эффект альтернации» [8, 18, 23, 7, 14, 24]. Если объекты одного таксономического ранга выстроены в порядке эволюции, то альтернируют их внешние свойства – фенотип. Различаются три типа альтернации. По суммарному объёму объектов в таксонах (табл. 1), по разнообразию объектов в таксонах (табл. 2) и по качественным или фенотипическим (внешним) признакам объектов (табл. 3).
Таблица 1
Тип 1. Альтернация по суммарному объёму объектов в таксонах

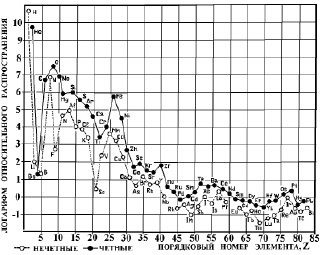
Рис. 1. Космическая распространенность химических элементов [8, 7]
Таблица 2
Тип 2. Альтернация по разнообразию объектов в таксонах
|
АТОМЫ |
ЖИВЫЕ ОРГАНИЗМЫ |
|
Элементы с нечётными порядковыми номерами монотипные или представлены двумя стабильными изотопами (рис. 2) [8]. |
Аналогичное явление наблюдается по количеству видов, родов в таксонах одного итого же ранга в порядке предполагаемой эволюции организмов (табл. 3). |
|
Если рис. 2 представить как встречаемость элементов в зависимости от числа их стабильных изотопов, обнаружится зависимость Рис. 3. Монотипных элементов – 36 %, а вместе с двувидовыми – 54 %. |
В биологии обнаружена аналогичная зависимость (рис. 3) [16]. Монотипных родов более трети, а вместе с двувидовыми более половины. Эту зависимость называют «вогнутая кривая систематиков». |
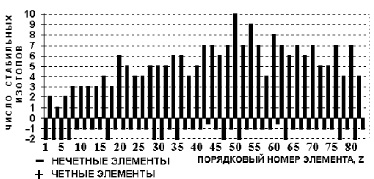
Рис. 2. Количество стабильных изотопов по элементам [8]
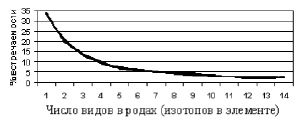
Рис. 3. Встречаемость таксонов с тем или иным разнообразием
К. Вилли, В. Детье расположили таксоны в порядке возрастания сложности [6]. Процесс эволюции, по их мнению, имеет ветвистый характер и поэтому они не смогли расположить все организмы в единый эволюционный ряд. По этой же публикации было подсчитано число видов, и обнаружилась альтернация второго типа (табл. 3).
Таблица 3
Обзор растительного и животного царств по К. Вилли, В. Детье [6]

Таблица 4
Альтернация по качественным или фенотипическим признакам
|
АТОМЫ, МОЛЕКУЛЫ |
ЖИВЫЕ ОРГАНИЗМЫ |
|
А. Как правило физико-химические свойства однотипных молекулярных веществ, выстроенных в порядке эволюции, альтернируют (Рис. 4.) [18]. |
А. Дивергенцию фенотипа между таксонами можно представить как эффект альтернации (рис. 5), так же как в химии (рис. 4). |
|
Б. Изотопы и элементы в зависимости от «чет-нечет» альтернативно различаются по физико-химическим свойствам [24]. В. В таблице Менделеева Д.И. у элементов в группах по вертикали, вероятно, также альтернируют физико-химические свойства в зависимости от чет–нечетности периодов. |
Б, В. С появлением новых видов или родов (таксонов) в порядке эволюции, их фенотип альтернирует, разбиваясь на 2 «подгруппы», которые могут «ошибочно» объединяться по фенотипу в разные таксоны или, наоборот, объединяются, хотя по генотипу это виды различных родов (таксонов), например, четных (рис. 5, 6, 7). Гомологично рис. 4. |
Рис. 4. Альтернация монокарбоновых и дикарбоновых кислот [18]
Альтернация может принимать форму дивергенции, параллелизма и конвергенции фенотипа на разном уровне таксонов (рис. 5).
В биологии приводят факты дивергенции таксонов по фенотипу, но точек ветвления, как правило, не находят. А альтернация фенотипа подразумевает, что точек ветвления просто нет и первые виды (таксоны) ближе к той или иной ветви по фенотипу (рис. 4, 5, 6, 7).

Рис. 5. Альтернация дивергенции, параллелизма и конвергенции таксонов
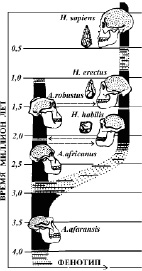
Рис. 6. Родословное дерево африканских гоминид [29]
Темный фон – время датированных находок, светлый фон – перерывы в геологической летописи. Четыре вида от A. africanus до H. erectus, соединенные автором стрелками в предполагаемой последовательности альтернативного возникновения и могут быть видами полирода. А виды A. afarensis и H. sapiens, могут принадлежать своим моно родам. Следует обратить внимание, что разрывы в датировках видов предполагаемого полирода относительно малы и равны, а между поли родом и монородами значительно больше.
Вверху (рис. 7) приведены две версии филогенетического дерева гоминид, построенные на основании сравнения их ДНК [1]. Внизу (рис. 7) представлена альтернация фенотипа таксонов большего порядка системности на основании эволюции гоминид в две мутации.
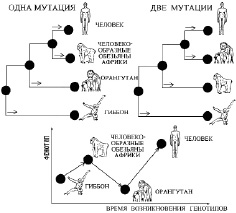
Рис. 7. Две версии филогенетического дерева гоминид [1]
Эволюция материи представлена рядом четных и нечетных таксонов; ЧАСТИЦЫ, АТОМЫ, МОЛЕКУЛЫ, ОРГАНИЗМЫ и т.д. Четные (или нечетные) таксоны должны быть более гомологичны между собой, чем рядом стоящие. На этом основании сопоставим АТОМЫ и ОРГАНИЗМЫ, как чётные таксоны и рассмотрим их систематики, как гомологичные признаки (табл. 5).
До Д.И. Менделеева элементы систематизировали по физико-химическим свойствам (фенотипу), составляли в триады, октавы, и другие гармонические группировки [31]. В биологии, опираясь на фенотип, систематики не могут прийти к общему мнению, даже по поводу царств живой природы (2 царства, 3, 4, 7, 9, 18), предлагают и другие всевозможные гармонические группировки и объединения.
Проведем мысленный опыт. Будем систематизировать атомы, опираясь на их физико-химические свойства и различия (фенотип). Добавим гомологичные предпосылки и положения, принятые в биологии. Будем считать, что атомы «рождаются» из «родственных» по фенотипу атомов. Объединяем атомы по внешним химическим свойствам в таксоны первого порядка изотопы–виды. «Близкородственные» по свойствам изотопы в элементы – таксоны второго порядка – роды. Проводим последующие объединения. Так или иначе, получатся иерархические таксоны рангом выше таксона – элемент. Если обратить внимание на горизонтальные, вертикальные и диагональные соответствия физико-химических свойств элементов, известные из периодической таблицы Д.И.Менделеева, то неизбежно получится «дерево» фенотипической эволюции атомов дивергентного типа (рис. 8), имеющуу соответствие и в биологии (рис. 9).
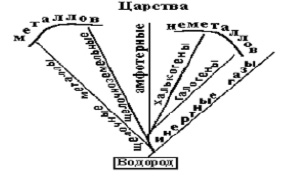
Рис. 8. Систематика элементов по внешним химико-физическим свойствам
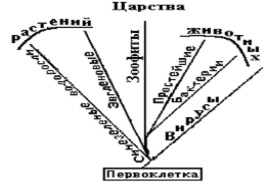
Рис. 9. Систематика живых организмов по фенотипу (внешним свойствам)
Периодический закон Д.И. Менделеева это естественная систематика, выстраивающая в ряд генотипические таксоны, а фенотипические периодично. Ю.А. Урманцев [35] утверждает, что: «Таблица Д.И. Менделеева – лишь первая или одна из первых эмпирически найденных реализаций периодической системы и в том числе не менделеевского типа, бесчисленное множество».
Согласно законам системности, явления альтернации и в соответствии с чётностью таксонов «АТОМЫ» и «ОРГАНИЗМЫ», в биологии должна быть естественная систематика гомологичная таблице Д.И. Менделеева (рис. 11), где таксон вид соответствуют изотопам, а роды – элементам. Для аналогий приводятся абстрактные схемы таблицы Д.И. Менделеева в биологии и в химии (рис. 10, 11) и рассматриваются их соответствия (табл. 5).

Рис. 10. Схема таблицы Менделеева Д.И. в химии
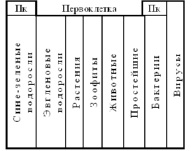
Рис. 11. Схема таблицы Менделеева Д.И. в биологии
Таблица 5
Соответствие признаков периодических таблиц живой и неживой природы
|
ПТМ АТОМОВ |
ПТМ ЖИВЫХ ОРГАНИЗМОВ |
|
1. Водород одновременно имеет свойства металлов и неметаллов. |
1. Первоклетка должна была иметь одновременно свойства растений и животных. |
|
2. В первых малых периодах свойства элементов относительно быстро сменяются от металлов к неметаллам. |
2. Первые периоды должны быть РНК–геномными одноклеточными организмами и их свойства должны сменяться от продуцентов к консументам. |
|
3. Инертные газы появились после водорода и замыкают все периоды, называются недеятельными элементами и этим свойством отличны от всех других элементов. |
3. Вирусы появились после Первоклетки и замыкают периоды. Отличны от всех остальных организмов и считаются реликтовыми формами жизни [37], как бы границей между живым и мертвым. |
|
4. Крайние группы – щелочноземельные и щелочные металлы, халькогены и галогены, по физико-химическим свойствам очень гомологично-консервативны. |
4. Крайние группы – сине-зеленые, эвгленовые водоросли, простейшие и бактерии фенотипически консервативны и на протяжении всей эволюции остаются одноклеточными. |
|
5. В периодах физико-химические свойства элементов все более приобретают неметаллические свойства вплоть до инертных газов. |
5. В периодах свойства организмов должны изменяться от продуцентов i-го порядка, переходящих в зоофиты, и далее в консументы i-го порядка до простейших, бактерий и вирусов. |
|
6. Горизонтальные, вертикальные и диагональные физико-химические свойства позволяют объединять элементы в «фенотипические» таксоны с эффектом параллелизма в высших таксонах, дивергенция, конвергенция, тупиковые линии и другие эффекты объединений в иерархические таксоны по фенотипу. |
6. Горизонтальные, вертикальные и диагональные фенотипические соответствия позволяют объединять таксон – род в различные иерархические таксоны. В биологии наблюдается параллелизм фенотипа высших таксонов [29], гомологично группам в химии. При этом эффект альтернации фенотипа или дивергенции должен проявляться за счет ступенчатого увеличения периодов и, соответственно, наблюдаться эффект тупиковых линий, и т.п. эффекты. |
|
7. Единый «генетический» материал – частицы. |
7. Единый генетический материал – триплеты. |
Не менее 7 соответствий указывают на существование периодического закона больших периодов и в биологии. Но нельзя предполагать, что периодический закон Д.И. Менделеева в химии точно повторяется в биологии. Так как биология иерархически выше, чем химия элементов и должна иметь дополнительные функции.
К настоящему времени накопилось свыше 160 вариантов различных представлений периодического закона Менделеева Д.И. [31]. Периодическую таблицу (рис. 11) можно представить в виде ступенчатой спирали (за счет увеличения периодов), и если ее разрезать на зоофитах (рис. 12), то середина таблицы (или время по периодам) будет представлена только одноклеточными микроорганизмами, что и вызывает прерывистое равновесие многоклеточных.
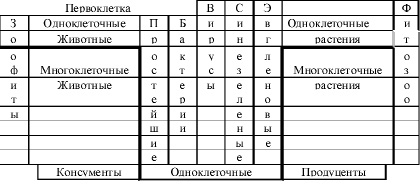
Рис. 12. Иное графическое представление таблицы Д.И. Менделеева в биологии
Периодически должно повышаться вида разнообразие как одноклеточных, так и многоклеточных таксонов, активность и суммарная численность одноклеточных микроорганизмов. Период таблицы рис. 11 заканчивается новым родом бактерий и вирусов, которые «сортируют и комбинируют» генетический материал всего предыдущего периода. Одновременно новые бактерии и вирусы, как патогенны, «освобождают» пищевые ниши для следующего периода, вызывая массовые вымирания многоклеточных живых организмов законченного периода, где катастрофические и космологические негативные факторы могут снижать иммунитет всех организмов к бактериально-вирусным инфекциям и ускорять процессы эволюции. Кювье назвал эти периодические явления «Мировыми катастрофами» [29]. В мезозое массовые вымирания наблюдались с периодичностью в 26, а в кайнозое 35–40 миллионов лет, вероятно из-за увеличения периодов, как и в химии. Периодические вымирания многоклеточных, совпадают с биогенным накоплением урана [21], за счёт активности в этот период одноклеточных. А.Л. Чижевским [41] обнаружены малые гармоники периодичности микроорганизмов, а должны быть и большие гармоники.
Гипотеза о периодическом законе Д.И. Менделеева в биологии объясняет, дополняет и согласуется с пятью основными закономерностями эволюции [6].
«1 закономерность – Эволюция происходит с разной скоростью в разные периоды. В настоящее время она протекает быстро, что выражается в появлении многих новых форм и вымирании многих старых».
«2 закономерность – Эволюция организмов различных типов происходит с разной скоростью. Вообще эволюция протекает быстрее при первом появлении нового вида, а затем, по мере стабилизации группы, постепенно замедляется».
Появление новых вирусов и бактерий определяют скорость появления новых видов многоклеточных нового периода и последних циклов любых меньших гармоник в отличии от оставшихся старых видов предыдущих периодов или меньших циклов.
«3 – Новые виды образуются не из самых высокоразвитых и специализированных форм, а, напротив, из относительно простых, неспециализированных».
«4 закономерность – Эволюция не всегда идет от простого к сложному. Существует много примеров «регрессивной» эволюции, когда сложная форма давала начало более простой».
Высокоразвитость, специализированность, многообразие, однаклеточность и многоклеточность – это признаки полигенного фенотипа таксонов, который должны периодически альтернировать. К примеру, периодический закон в биологии предполагает, что по большему периоду за фенотипически «примитивными» одноклеточными водорослями в естественной систематике появляются «высокоразвитые» многоклеточные растения, затем фенотипически более «примитивные» зоофиты; после «высокоразвитые» многоклеточные животные, а затем – простейшие одноклеточные и вирусы. То есть, дважды за один большой период относительно «простые» по фенотипу организмы образуются из высокоразвитых по фенотипу организмов. То же самое гомологично должно происходить и по меньшим гармоникам альтернации фенотипа.
«5 закономерность – Эволюция затрагивает популяцию, а не отдельные особи и происходит в результате процессов мутирования, естественного отбора и дрейфа генов».
Эволюция в биологии – это системное изменение генофонда от вида к виду, от рода к роду, которое происходит за счет трансдукции вирусов и затрагивает не просто популяцию отдельного вида, а весь подготовленный фоновыми мутациями генофонд живой материи в основном настоящего и предыдущего периодов. Наследственно новые виды и роды организмов по средству вирусов могут появляться из видов и родов, расположенных горизонтально, вертикально и диагонально в таблице Д.И. Менделеева.
В средние века наблюдалась периодическая активность бактерий животных и человека. В то же время, должны были быть гомологичные явления с вирусами у растений. В текущий момент времени наблюдается активность вирусов у животных, а в растительном мире массовая гибель древесных пород от бактериозов [44, 39]. Затем волна бактериозов растений перейдет на животных, а волна вирусных болезней животных затухнет, и начнется вирусная волна у растений, и так далее, периодически сменяя друг друга.
На основании периодической системы можно найти фундаментальные признаки времени в биологии, которые позволят определить последовательность появления генотипов. Фундаментальные признаки времени в биологии должны быть гомологичны химическим – массе, размерам атомов и т.п.. Л.Л. Численко [42] отметил, что численные значения средних размеров особей различных биологических таксонов в логнормальной декартовой системе координат отстоят друг от друга в среднем на 50 логарифмических единиц. Что соответствует значению Пи с точностью до третьего знака после запятой. С.И.Сухонос [32] гомологично показал, что размеры одного и того же порядка системности отстоят друг от друга в логнормальной декартовой системе координат на определённый интервал размерности кратный значению 2Пи (рис. 13).
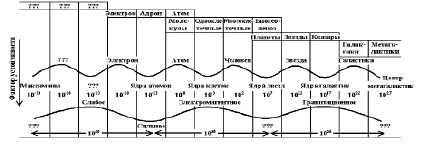
Рис. 13. Размеры классов или объект-систем в десятичных логарифмах [32]
Все материальные объекты периодически занимают масштабные уровни эволюции, подразделяясь на два альтернативных таксона, ядерный и «неядерный». Свойства (фенотип) «неядерных» объектов определяются внутренней (генетической) структурой ядерных объектов; ядра атомов определяют свойства атомов, ядра клетки – свойства организмов и т.д. Эта взаимосвязь таксонов и их различная относительная устойчивость позволяет классифицировать объекты в виде волновой модели. Четные на гребне – неустойчивое равновесие, нечетные во впадине – устойчивое. Ось волны в местах пересечения дает достаточно точное значение размеров основных классов объектов. На волне с масштабом в 1020 объект-системы с жестко детерминированными внутренними связями принадлежат левым склонам нечетных волн, вплоть до их гребней (атомы, планеты, звезды), а на правых склонах усложняются, группируются в структуры (молекулы, субклеточные, тесные и широкие звездные пары, планетные системы), вплоть до начала нечетных волн.
Периодичность полигенных признаков популяции и индивидуума…
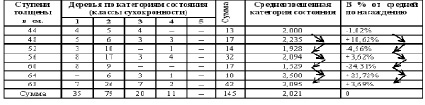
В 1981 году при статистической обработке полевых материалов лесопатологического обследования Тульских засек, отмечена синхронная периодичность сухокронности дуба черешчатого по когортам поколения (табл. 6, 7).
Сухокронность – признак реакции дерева (иммунитет) на отрицательные факторы среды (бактериоз) и определяется категорией состояния по проценту сухих ветвей в кроне. Ступень диаметра дерев – это функция времени (возрастная когорта поколений).
Таблица 6
Альтернация сухокронноти по когортам поколений (Яснополянское лесничество квартал 23 выдел 21)
На постоянной пробной площади 2 (табл. 2) наблюдается аналогичная альтернация сухокронности, но здесь представлена динамика между 1980 годом и 1981 годом. Если категория сухокронности между учетами не изменилась, ставился балл 0, если происходило ухудшение, ставился балл равный изменению категории со знаком минус, если улучшение – то плюс соответствующий балл. Усохшие деревья при первом перечете не учитывались. Улучшение кроны, вызвано тем, что процесс усыхания кроны прекратился, а мелкие веточки кроны сгнивают и опадают из-за ветра и снега.
Таблица 7
Альтернация сухокронноти по когортам поколений (Проба 2 Музей «Ясная поляна», Арковский верх)
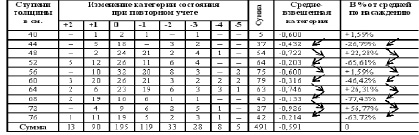
Категория, в зависимости от четности-нечетности, хуже у 40 – 48 – 56 – 64 – 72 ступеней толщины и лучше у 44 – 52 – 60 – 68 – 76 ступеней толщины. Ступень толщины, случайно совпадающая на момент измерения с определенными естественными возрастными когортами деревьев. Альтернация признака синхронности на множестве пробных площадей подразумевает наличие гармоники периодичности на уровне всей популяции дуба. Цикл периодичности равен 4 см + 4 см = 8 см. Средний годичный прирост дуба по спилам в среднем равен 2,8 мм, что согласуется с таблицами хода роста [12]. Соответственно гармоника в реальном времени примерно равна 8см / 2,8 мм ≈ 29 годам, а полупериод или когорта альтернации приблизительно 14,5 годам (4 см).
Радиальный прирост деревьев подчиняется логистической кривой (возрастной тренд), поэтому данные преобразовывались в кумулятиву, тренд которой описывался полиномиальной функцией. Затем эта функция преобразовывалась в не кумулятивную функцию, которая и вычленялась из исходного ряда данных, образуя остатки. По остаткам и их усредненным рядам нечетными средне скользящими (3, 5 и т.д.) каждый раз производился автокорреляцией анализ с отбором кривых автокорреляции, которые наибольшим образом отражали регулярность гармоники периодичности. Чем больше средне скользящая осреднения, тем меньше проявление «Белого статистического шума» [43], зависящего от количества элементов в группах выборки. Группами выборки в этом случае являются средне скользящие, состоящие из множества элементов.
Для анализа прочих параметров периодичности должен применяться цифровой спектральный анализа [26 19], но наши данные заведомо искажают амплитуду признака систематическими и случайными погрешностями прироста, так как возраст у нас определялся по косвенному признаку – диаметр или периметр деревьев. Поэтому данную работу надо рассматривать как предварительную, и эти явления следует проанализировать по реальному возрасту деревьев на вырубках или по картотекам поликлиник для человека.
Периодичность выявлена у 5 древесных пород (дуб, липа, кедр, пихта, ель) и всем проанализированным 42 лесопатологическим и фенотипическим признакам [26, 27, 28].
В 1993 году в Тульских засеках была заложена сеть постоянных пунктов учета лесопатологического мониторинга. Повторные учеты проводились в 1994 и 1997 годах. Все данные мониторинга были объединены в один массив данных по породам. Вместо диаметра измерялся периметр дерева на высоте 1,3 м. Среднегодичный прирост деревьев второго бонитета равен 2,8 мм [12] или 8,8 мм периметра.
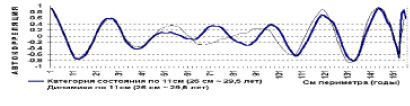
Рис. 14. Дуб. Автокорреляция категории состояния кроны и ее динамики
При средне скользящей в 11 см периметра обнаруживается гармоника в 26 см периметра или 8,3 см диаметра, что соответствует 29,5 годам, а полупериод 14,8 годам или 13 см периметра (4,1 см диаметра). Аналогичная периодичность и была независимо обнаружен в 1981 году на территории музея «Ясная поляна» и в Тульских засеках при ступенях толщины в 4 см. Это периодичность (альтернация) 3 типа.
В 1984 году в Бурятской АССР в кедрово-пихтовом насаждении была заложена большая пробная площадь.
Э.Н. Фалалеев [36] отмечает, что в процессе роста пихтарников, запас древесины на 1 га строго периодичен (рис. 16) и соответствует рис. 15 по количеству деревьев на выделе.
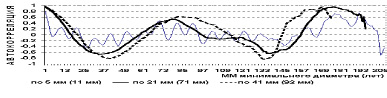
Рис. 15. Автокорреляция количества деревьев пихты по периметрам
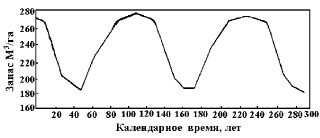
Рис. 16. Периодическое изменение древесного запаса в пихтарниках [36]
Это периодичность (альтернация) 1 типа. Все проанализированные фенотипические признаки периодичны по когортам поколений и отражают скорость и объем их биохимического метаболизма. У человека это отражает работа печени. Выявлено, что скорость работы печени имеет двуглавую зависимость [2, 3]. Это может быть результатом необъективного объединения в выборку четных и нечетных когорт поколений, имеющих разные генеральные совокупности (Гаусс).
Генетическая среда состоит из простых элементарных генов и сложных – опероны и комплексы оперонов и последние могут быть более или менее активными. «Продуктами» генов являются гормоны или другие биохимические вещества, определяющие признаки фенотипа. Априорно между конкретными «продуктами» генов наблюдается функциональная симметрия, а значит и между генами. Каждому гормону или иному генетически синтезируемому веществу А (оперонам А или их комплексу) функционально симметричен антигормон или другой биохимический механизм компенсации Б (оперон Б или их комплекс), что надо рассматривать как целый объект-квант взаимодействия общей теории циклов (ОТЦ) [30]. Мужскому – женский гормон, гормону роста – антигормон или иной механизм биохимической компенсации, в противном же случае скорость, рост или значение другого конкретного полигенного признака был бы постоянен и неограничен при жизни организма, как и продолжительность жизни как полигенный признак. Вероятно, у четных (или нечетных) когорт поколений определенные опероны или их комплексы пассивны, а симметричные более активны (доминируют), а у последующих когорт наоборот. Этим объясняется механизм периодичности и различие гармоник у разных полигенных признаков, так как они определяются своими комплексами оперонов разного размера и последовательности, а значит и своими часовыми механизмами периодичности в структуре ДНК генотипа индивидуума, генофонда популяции, вида или всей живой материи.
Часовые механизмы могут быть эндогенными и навязанные внешними периодическими факторами [22, 47]. Некоторые исследователи связывают часовые механизмы биоритма непосредственно с ДНК [11]. По мнению автора, внешние факторы могут влиять только на амплитуду цикла, а эндогенные механизмы определяют его период.
А.Л. Чижевским [41] отмечена периодичность в популяциях микро и макро организмов. С.С. Четвериков [40] определил подобные явления как «волны жизни». Этим же явлением объясняются синхронные, модные мутации в популяциях дрозофил [15, 13], модные мутации видов в роде, между родами в семействах и далеко отстоящих таксонов [9].
Дарвиновский отбор [46], как внешний факторов среды, может разрешить или запретить полигенный признак, но создать тот или иной полигенный признак одновременно у множества видов и родов, могут только эндогенные механизмы эволюции. Внешний фактор может запретить, ускорить или замедлить эволюцию, но не способен вызвать ее.
Зигфрид Коллер [45] утверждал, что следующие друг за другом поколения новорожденных, не обладают и не могут обладать одинаковой жизненной силой, есть более жизнеспособные когорты поколений и менее жизнеспособные. Популяционной периодичностью объясняются не детерминация результатов повторных опытов в биологии при поиске вещества памяти [46, 25].
С.Ю. Маслов [20] утверждал, что доминирование левого или правого полушария мозга в популяциях человека различно в различные исторические эпохи и иллюстрировал это сменой архитектурных стилей, господствовавших в Европе. В математике обнаруживал периоды, когда ведущую роль играла строгая логика, но в иные периоды возникало время интуиции, гениальных догадок и эвристических рассуждений. Л.Н. Гумилев [10] писал, об этом же явлении, но о гармониках с большим периодом. Этим же объясняется смена реалистичных наскальных рисунков периода палеолита схематично абстрактными рисунками в неолите (еще большие гармоники биоритма).
По полу зачатого ребенка выявлена периодичность физиологического состояния среднестатистического мужчины. Вместо этого признака можно подставить любой другой биометрический или альтернативный признак-симптом физиологии или симптом болезни.
Теоретически соотношение полов определяется фактором случайности. Но на 100 новорожденных девочек приходится в среднем 107 мальчиков и нормой для человека считается половой индекс 1,07 [38, 34, 33]. При этом частота рождения мальчиков максимальна при первых родах и падает при последующих. В семьях долгожителей высокая частота рождения мальчиков; у больных подагрой преобладают девочки; в потомстве лысых мужчин в полтора раза больше мальчиков; есть связь с комплекцией, темпераментом родителей и с их профессией. Следовательно, у Х и Y сперматозоидов оплодотворить яйцеклетку и зависит от физиологических и патофизиологических особенностей индивидуума.
Период выживания сперматозоидов 1,5 – 72 часа. Y–сперматозоиды в среднем мельче и более подвижны – меньше сопротивление в жидкой среде, чем и объясняется преобладание рождений мальчиков. Но Y–сперматозоиды менее жизнеспособны по сравнению Х–сперматозоидами. Следовательно, если во всех сперматозоидах достаточное количество цитоплазмы для преодоления пути, то должны добираться первыми до яйцеклетки Y–сперматозоиды и рождаться мальчики. Если же в целом у Y–сперматозоидов недостаточное количество цитоплазмы, то должны приходить к яйцеклетке Х–сперматозоиды и рождаться девочки.
Статистический материал (табл. 8) собирался в 1984-85 годах в детских садах. Учитывалась дата (месяц) рождения отца, дата (месяц) рождения и пол ребенка. От месяца рождения отца отсчитывался месяц рождения ребенка, тем самым анализировался только средний годовой цикл от момента рождения отца. Например, отец родился в апреле, а ребенок в ноябре, значит, ребенок родился через восемь месяцев относительно рождения отца по его годовому циклу.
Таблица 8
Соотношение мальчиков и девочек в детских садах (годовой цикл)
|
Пол ребенка |
Месяцы рождения ребенка, после месяца рождения отца |
Сумма |
|||||||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
||
|
Мальчик |
20 |
14 |
19 |
19 |
16 |
16 |
4 |
7 |
1 |
6 |
5 |
4 |
131 |
|
Девочка |
8 |
6 |
10 |
4 |
10 |
9 |
11 |
9 |
12 |
15 |
19 |
8 |
121 |
|
В сумме |
28 |
20 |
29 |
23 |
26 |
25 |
15 |
16 |
13 |
21 |
24 |
12 |
252 |
|
Половой индекс |
2,5 |
2,33 |
1,9 |
4,75 |
1,6 |
1,78 |
0,36 |
0,78 |
0,08 |
0,4 |
0,26 |
0,5 |
1,08 |
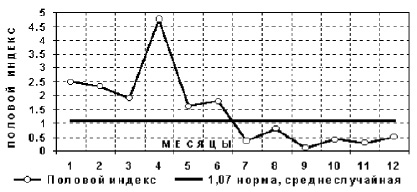
Рис. 17. Половой индекс в детских садах по годовому циклу отца
Средний половой индекс выборки (1,08) и хорошо согласуется со среднестатистическими данными (1,07). Из табл. 8 и рис. 17 видно, что в первом полугодии после месяца рождения отца значительно преобладают рождения мальчиков относительно девочек, а во втором полугодии, наоборот, девочек рождается больше чем мальчиков.
Рис. 18 отражает относительный процент рождения мальчиков и девочек в каждом месяце (сумма детей в каждом месяце принята за 100 %).
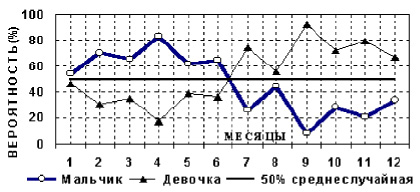
Рис. 18. Относительный процент рождения мальчиков (девочек) по годовому циклу отца
На рис. 18, наряду с годовой периодичностью отражена альтернация кривой в зависимости от четности и нечетности месяцев. Вероятно, это 28-дневный цикл, гомологичный женскому. Как и у биоритма популяции, у индивидуума должны быть и другие гармоники периодичности, например, четные – нечетные или кризисные годы.
Методически анализ построен по принципу периодической таблицы Менделеева Д.И., только все усреднено в один период равный году. Но годовой цикл может быть чуть больше или меньше и значит, может вкрасться систематическая ошибка сглаживания амплитуды цикла. Маловероятно, что месячный цикл кратен годовому циклу, и, тем самым тоже сглажен. Кроме того, не учитывался фактический срок беременности. Ведь часто мальчики и отцы недоношены, а девочки переношены.
Выявлено что если совокупление совершалось до овуляции, вероятность рождения девочки 75-80 %, а если оно произойдет сразу после овуляции, то на 80-90 % родится мальчик [5]. Естественно, если совокупление происходит до овуляции, Y–сперматозоиды просто не выживают к моменту появления яйцеклетки, и в этом случае рождаются девочки. А если совокупление происходит после овуляции, то Y-сперматозоиды быстрее достигают яйцеклетки и рождаются мальчики.
Если учесть все вышеизложенное, изучить возрастные гармоники индивидуума и гармоники когорт популяции, а также учесть влияние факторов среды, то точность и надежность прогноза может значительно увеличиться.
Y-сперматозоиды в среднем мельче, чем Х, но если априорно рассматривать только Y (или Х) сперматозоиды то, среди них должны встречаться как очень мелкие, так и наиболее крупные согласно закону Нормального распределения (Гаусс), для каждого типа сперматозоидов отдельно рис. 19.
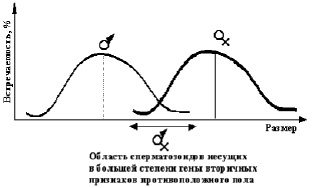
Рис. 19. Априорное распределение сперматозоидов по типам в зависимости от их размеров
Возможно, чем больше или меньше размер Y-сперматозоид (или Х), тем он больше несет цитоплазменных гормонально эндогенных механизмов определенным образом запускающих ядерные гены, ответственные за вторичные половые признаки противоположного пола. Если это так, то должны с определенной вероятностью по биоритму распределяться рождения гермафродитов, а также предрасположенных к однополым связям мужчин и женщин.
Зная текущие периоды гармоник трех и более признаков полигенного фенотипа поколений популяции, можно определить возраст таксона по сопоставлению гармоник с началом автокорреляции – полным их резонансом. Этот метод определения возраста таксонов можно проверить на практике по индивидууму человека с известным моментом зачатья (рождения).
Выявленные периодические явления в биологии и составление по ним графиков и таблиц периодичности фенотипических признаков индивидуума, поколений популяций, видов, родов и всей живой материи, позволит разобраться в структуре генов живой материи и полигенной наследственности. Подобно тому, как это теоретически определено в ядерной физике, опираясь на периодическую таблицу Д.И. Менделеева. Карты и графики биоритмов индивидуума и популяции найдут применение в медицине для прогнозов заболеваний для индивидуума, эпидемий для поколений популяции и выявления детерминированной реакции организмов на факторы среды, определения прогноза эффективности тех или иных доз лекарств и методов лечения индивидуума, а также в селекции растений и животных.
Если таксоны одного масштаба системности выстроить в порядке эволюции их генотипов во времени, то наблюдается расслоение таксонов одного ранга на две качественно различные по фенотипу симметричные подгруппы: четные и нечетные табл. 9.
Таблица 9
Девять уровней альтернации или периодичности фенотипа объектов

Краткая формулировка выявленного Всеобщего периодического закона: таксоны разного ранга системности, выстроенные в порядке их появления в эволюции, периодически изменяют значения своих полигенных признаков по независимым гармоникам периодичности, определяемым эндогенными причинами самих этих объектов и всего мироздания.
Библиографическая ссылка
Сафронов В.Н. ПЕРИОДИЧЕСКАЯ ЭВОЛЮЦИЯ В БИОЛОГИИ И ДРУГИХ ОБЪЕКТ СИСТЕМАХ // Международный журнал прикладных и фундаментальных исследований. 2014. № 9-1. С. 124-139;URL: https://applied-research.ru/ru/article/view?id=5803 (дата обращения: 02.08.2026).