

В контексте проблемы глобального изменения климата одной из актуальных является проблема устойчивости гидрологического цикла в лесных экосистемах. Для оценки устойчивости сообществ и слагающих их видов к колебаниям климата необходимо располагать количественными характеристиками основных показателей состояния растений, которые можно рассматривать как норму для данных условий произрастания. Анализ известных данных [9, 8, 6, и др.] показывает как различия показателей водного обмена у разных видов в одинаковых экологических условиях, так и их сходство у растений одного вида, но из разных мест обитания. Это свидетельствует о видоспецифичном характере процесса водного обмена, и, казалось бы, позволяет привлекать для анализа данные, полученные разными авторами. Однако неадекватность методик и длительности исследований порой дают значительные различия в результатах.
Целью нашей работы было сравнительное исследование закономерностей варьирования переменных водного режима хвойного (Pinus sylvestris L.) и лиственного (Betula pendula Roth) видов растений в суточной, сезонной и межгодовой динамике при их совместном произрастании в условиях достаточной почвенной влагообеспеченности.
Материалы и методы исследования
Исследования проводили в сосняке черничном свежем европейской части средней тайги в Южной Карелии (N 62о13′, Е 34о10′). Таксационное описание древостоя и характеристика почв представлены в работе [4]. Объектами исследования послужили 20-летние деревья сосны обыкновенной (Pinus sylvestris L.) и березы повислой (Betula pendula Roth). Сопряженные исследования показателей водного режима и запасов влаги в почве проводили с мая по сентябрь в течение вегетационных периодов 1977–1997 и 2013 гг. Наблюдения проводили для 10 деревьев; с каждого дерева отбирали по 3 побега со средней части кроны. Водные потенциалы почвы определяли с помощью тензиометра [7], водные потенциалы корней (Ψк) и облиственных (охвоенных) побегов (Ψпоб.) – с помощью камеры давления Plant Moisture Vessel SKPM 1400 (Skye Instruments Ltd., Великобритания). Для измерения скорости водного потока применяли тепловые методы [3], в частности, для определения относительной скорости (Vотн.) – метод стационарного нагрева, для абсолютной скорости (Vабс.) – импульсный метод. Метеорологические параметры определяли стандартными метеоприборами и с помощью системы LI-COR 6400ХТ (LI-COR Inc., США). Для обработки экспериментальных данных использовали методы вариационной статистики. Проверку гипотез и оценку существенных различий между средними величинами осуществляли с помощью критерия Стьюдента при 5 %-ном уровне значимости.
Результаты исследования и их обсуждение
Сравнительный анализ переменных водного режима хвойного и лиственного видов показал, что водные потенциалы корней (Ψк) деревьев сосны и березы были близки по величине и размаху колебаний в течение суток. Так, для сосны средние за вегетацию предрассветные и дневные значения Ψк составили – 0,22 ± 0,02 и – 0,32 ± 0,03 МПа, для березы – – 0,21 ± 0,03 и – 0,27 ± 0,04 МПа, соответственно. Следует отметить, что водные потенциалы почвы (Ψп) в сосняке черничном свежем в зоне наибольшего распространения корней имели высокие значения в течение ряда вегетационных периодов с мая по сентябрь: значения Ψп не опускались ниже – 0.05 МПа, что свидетельствовало о достаточной влагообеспеченности растений. Наряду с этим, вследствие мозаичности влажности почвы в пределах объема корневой системы дерева, для исследуемого показателя Ψк была отмечена определенная пространственная вариабельность, которая у сосны была ниже по сравнению с березой. Так, например, коэффициент вариации при определении средних значений Ψк для сосны и березы составил 13–15 и 15–25 %, соответственно.
Сравнительное исследование водных потенциалов охвоенных (облиственных) побегов (Ψпоб.) сосны и березы также показало сходство их суточной динамики. При этом следует отметить более высокие предрассветные величины Ψпоб max березы по сравнению с сосной. Так, например, максимальные предрассветные Ψпоб max березы и сосны составили – 0,1 и – 0,3 МПа, соответственно. Однако сопоставление минимальных значений дневных водных потенциалов охвоенных (облиственных) побегов (Ψпоб min) сосны и березы, в отличие от максимальных значений предрассветного показателя, выявило разные варианты. Так, в первом случае дневные значения Ψпоб min, также как и предрассветные значения Ψпоб max, были выше у березы по сравнению с сосной; во втором случае, напротив, дневные значения Ψпоб min березы были ниже, чем у сосны; и в третьем – Ψпоб min обоих видов были схожими. Первый вариант наблюдали в начале вегетации (июнь) в фазу распускания листьев у растений березы, второй – в середине вегетации (июль) в фазу появления и роста молодой хвои у растений сосны. Эти периоды были довольно короткими, и большую часть вегетации наблюдали близость значений Ψпоб min обоих видов. Так, наименьшие дневные значения показателя Ψпоб min составили для сосны и березы – 1,5 и – 1,6 МПа, соответственно.
Изменения Ψпоб в течение суток были связаны, прежде всего, с колебаниями погодных условий, поскольку, как показано выше, почвенная влагообеспеченность растений была достаточной. Диапазоны изменчивости Ψпоб max и Ψпоб min сохранялись в разные вегетационные периоды, что явилось основанием для расчета их средних за вегетацию величин. В разные годы наблюдений значения предрассветного и дневного показателя варьировали в незначительных пределах и составили для сосны – 0,44 ± 0,07 … – 0,43 ± 0,05 и – 1,01 ± 0,14 … – 0,97 ± ± 0,07 МПа, для березы – соответственно, – 0,24 ± 0,02… – 0,22 ± 0,02 и – 1,14 ± 0,08 …– 1,0 ± 0,02 МПа. При сравнении максимальных и минимальных значений Ψпоб в разные годы исследования значимых различий не обнаружено (р < 0,05), что свидетельствует о постоянстве показателей в межгодовой динамике.
Наблюдаемые различия в величинах Ψк и Ψпоб у сосны и березы, а также рассчитанные по этим данным градиенты водных потенциалов (grad Ψ = (Ψпоб. – Ψк)h) – движущие водные потоки силы, были не столь велики, как полученные различия в скорости потоков влаги по стволам этих видов. Так, максимальные значения линейной (V) и объемной (U) скорости, регистрируемые в летние месяцы, составили у березы и сосны, соответственно, 26,5 см∙час-1 и 13,81 г∙см-2∙час-1, 15,5 см∙час-1 и 9,95 г× ×см-2∙час-1. Средние за вегетационный период среднесуточные скорости V и U у сосны были 4,6 см∙час-1 и 3,2 г∙см-2∙час-1, а у березы – 8,68 см ч-1 и 5,65 г см-2 ч-1, соответственно. Этот факт свидетельствует о более высоких скоростях водных потоков у растений березы по сравнению с сосной. Вероятно, основная причина этих различий связана, прежде всего, с особенностями строения древесины сосны и березы. Известно, что основными элементами древесины хвойных являются трахеиды, которые представляют собой мертвые клетки длиной от 0,5 до 15 мм. Стенки соседних трахеид содержат многочисленные поры, способствующие движению воды и растворов между трахеидами. Характерной чертой древесины покрытосеменных является наличие сосудов, состоящих из серии вертикальных клеток, у которых нет поперечной стенки [10]. Для березы, в частности, отмечают рассеянно-сосудистое строение древесины с сосудами до 10 см длиной и диаметром 20 мкм [1]. Вследствие этого, меньшие размеры трахеид у хвойных, по сравнению с размерами сосудов лиственных пород, являются причиной большего сопротивления движению воды у сосны по сравнению с березой и, как следствие, меньшей скоростью водного потока.


Рис. 1. Зависимость относительной скорости водного потока (Vотн) по ксилеме ствола от водного потенциала побегов (Ψпоб.) сосны (а) и березы (б)
Одним из косвенных доказательств более высокой проводимости ксилемы березы по сравнению с сосной послужили результаты анализа взаимосвязи между скоростью водного потока и движущих водный поток сил – водных потенциалов (рис. 1). Так, сопоставление относительной скорости водного потока (Vотн) и Ψпоб. показало, что увеличение движущей водный поток силы (уменьшение Ψ поб.) у сосны и березы до – 0,6 и – 0,4 МПа, соответственно, приводит к увеличению Vотн.. Далее изменения Ψ поб в определенном диапазоне, который составил для сосны и березы – 0,6…– 0,85 и – 0,4…– 0,6 МПа, соответственно, не влияют на Vотн. При снижении значений Ψ поб у сосны и березы более – 0,85 и – 0,6 МПа, соответственно, Vотн. уменьшается. Уменьшение водного потенциала характеризует, с одной стороны, увеличение движущей водный поток силы, что приводит к увеличению скорости потока, с другой – свидетельствует о нарастании водного дефицита в проводящей системе дерева. Ранее нами [5] было показано, что водный потенциал, соответствующий максимуму Vотн., является показателем начала изменения оводненности водопроводящей системы дерева. Таким образом, начало уменьшения оводненности ствола у березы происходит при более высоком значении Ψ поб, поскольку диапазон Ψпоб, которому соответствует максимальная скорость водного потока у березы (– 0,4… – 0,6 МПа), сдвинут в сторону более высоких значений по сравнению с сосной (– 0,6…– 0,85 МПа).
Другим свидетельством более высокого сопротивления движению влаги у сосны, по сравнению с березой, послужил анализ результатов регистрации переменных водного режима в ночное время. Как следует из наших данных, в звене «почва-растение» существует постоянный gradΨ , поскольку, как показано выше, Ψпоб ниже, чем Ψк. Поэтому, не смотря на уменьшение интенсивности транспирации в вечерние часы и прекращение ее при нулевой интенсивности ФАР [2], водные потоки, благодаря наличию градиента водного потенциала в звене «почва-растение», не прекращаются и в ночное время. В течение ночи скорость водного потока уменьшается, а водный потенциал увеличивается, достигая предельных за сутки величин к предрассветному часу. Взаимосвязь между Vотн.min и Ψmax аппроксимировали линейными уравнениями (рис. 2).
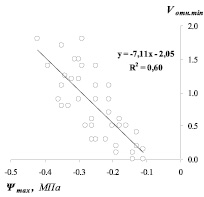
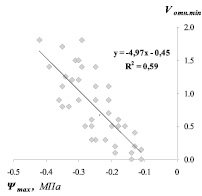
Рис. 2. Зависимость предрассветных значений относительной скорости водного потока (Vотн.min) по ксилеме ствола от водного потенциала побегов (Ψmax) сосны (а) и березы (б)
Vотн.min = – 7,11 Ψmax – 2,05 R2 = 0,60 (Pinus sylvestris L.)
Vотн.min = – 4,97 Ψmax – 0,45 R2 = 0,59 (Betula pendula Roth)
Более низкий (по абсолютному значению) коэффициент, показывающий угол наклона прямой, в уравнении связи для березы (– 4,97) по сравнению с сосной (– 7,11) является еще одним свидетельством более низкого сопротивления движению влаги у березы по сравнению с сосной.
Заключение
В условиях достаточного почвенного увлажнения нами выявлены диапазоны изменчивости параметров водообмена деревьев сосны и березы в течение ряда вегетационных периодов. Кроме того, установлена относительная стабилизация их в межгодовой динамике, что, в свою очередь, свидетельствует о произрастании исследуемых растений в пределах диапазона условий, где работают механизмы гомеостатирования. Выявленные отличия в большей степени обусловлены различиями в строении водопроводящих систем хвойного и лиственного видов. Однако одинаковый характер динамики показателей водного обмена сосны и березы может быть обусловлен сходством эволюционно выработанных адаптаций к меняющимся условиям среды.
Работа выполнена в рамках государственного задания ИЛ КарНЦ РАН и при частичной финансовой поддержке РФФИ (грант 13-04-00827-а).
Библиографическая ссылка
Сазонова Т.А., Софронова И.Н., Новичонок Е.В., Придача В.Б. ВОДНЫЙ РЕЖИМ ДРЕВЕСНЫХ РАСТЕНИЙ В УСЛОВИЯХ ДОСТАТОЧНОГО ПОЧВЕННОГО УВЛАЖНЕНИЯ НА СЕВЕРО-ЗАПАДЕ РОССИИ // Международный журнал прикладных и фундаментальных исследований. 2015. № 8-2. С. 299-302;URL: https://applied-research.ru/ru/article/view?id=7089 (дата обращения: 02.08.2026).