

В настоящее время большое внимание исследователей направлено на выявление механизмов, позволяющих растениям адаптироваться к изменению условий окружающей среды. Одним из наиболее важных абиотических факторов, влияющих на рост и развитие растений, а также поддерживающим структуру растительных сообществ, является световой фактор. Однако в лесных сообществах в результате различных естественных (вывалы крупных деревьев первого яруса) и антропогенных процессов (выборочные рубки леса) происходит образование лесных «окон», в которых наблюдается резкое изменение режима освещенности. Это приводит к тому, что растения должны достаточно быстро адаптироваться к изменению условий освещенности. В случае неспособности растений адаптироваться к увеличению повышенной освещенности возможно изменению структуры и видового состава сообщества. Особенно актуальна эта проблема в муссонных тропических лесах, так как в них растения подлеска и подроста эволюционно адаптированы к условиям очень низкой освещенности. Одной из важнейших характеристик, позволяющих растениям выживать в изменившихся условиях, является адаптация фотосинтетического аппарата. Способность к адаптации, которая позволяет видам растений реагировать на различия в освещенности, может быть определена как фенотипическая пластичность [6]. Несмотря на наличие большого количества работ, до настоящего времени нет четкого понимания роли фенотипической пластичности отдельных видов в сукцессионных процессах и при адаптации растений к факторам среды [2, 9]. Изменения в проявлении признаков в ответ на изменение условий среды могут повлиять на взаимодействие растений. Несмотря на то, что эволюционные аспекты фенотипической пластичности были хорошо исследованы, до сих пор недостаточно данных о влиянии пластичности на экологические взаимодействия между растениями [3]. Таким образом, анализ изменений фотосинтетического аппарата в ответ на изменение освещенности и оценка степени фенотипической пластичности может помочь лучше понять закономерности распределения видов в сообществе и оценить их способность участвовать в восстановительных сукцессионных процессах после нарушений целостности лесного полога.
В связи с этим целью нашей работы являлось определение степени фенотипической пластичности к свету морфо-биохимических показателей фотосинтетического аппарата растений подлеска и подроста муссонного тропического леса и их способности учавствовать в сукцессионных процессах после образования лесных «окон».
Материалы и методы исследования
Исследование было проведено в национальном парке Кат Тьен, расположенном на юге Вьетнама (11 °21’–11 °48’ с. ш.; 107 °10’–107 °34’ в.д.). Климат южного Вьетнама является тропическо-муссонным. Среднегодовые температуры воздуха составляют 26–27 °С, годовое поступление осадков – около 2450 мм, в течение года четко выделяются два сезона: сухой (ноябрь – апрель) и влажный (май – октябрь) [1].
Объектами исследования были 16 видов древесных растений, которые занимают разное место в высотном градиенте сообщества:
– деревья верхнего древесного подъяруса: Lagerstroemia calyculata, Dipterocarpus alatus;
– деревья среднего древесного подъяруса (2–3 подъярусов): Hibiscus macrophyllus, Dalbergia mammosa, Grewia paniculata, Pterospermum sp., Ochrocarpus siamensis, Barringtonia cochinchinensis;
– древесные растения подлеска (нижний древесный подъярус): кустарники (Anaxagorea sp., Phyllanthus sp.) и дерево (Cleistanthus sp.);
– лианы: Ancistrocladus tectorius, Bauhinia sp., Smilax sp., Korthalsia sp., Calamus sp.
В период исследования все виды растений находились в подлеске в одинаковых экологических условиях роста и развития, но на разных этапах онтогенеза: если растения подлеска занимали свои типичные местообитания, то остальные виды были на ранних этапах развития и занимали соответствующую этому этапу экологическую нишу.
Для изучения степени пластичности параметров листовой пластинки для каждого вида было выбрано по 10 растений: 5 произрастающих в местах с низкой освещенностью (относительно ненарушенным лесным покровом, степень сомкнутости полога равнялась от 65 до 90 %) и 5 растений, произрастающих в местах с высокой освещенностью (участки леса с нарушенным лесным покровом, степень сомкнутости полога равнялась от 10 до 55 %). Пробы растительного материала отбирали в течение сухого сезона 2012‒2013 гг. Для определения содержания фотосинтетических пигментов было отобрано по 2 листа с каждого растения. Для определения морфологических показателей листовой пластинки (удельная листовая поверхность (SLA), содержание сухого вещества в листе (LDMC), длина (LL), ширина (LW), площадь (LA), степень вытянутости (LS) листа) и содержания элементов минерального питания (азот (N), фосфор (P), калий (K)) было собрано по 5 листьев с каждого растения. Длину и ширину листовой пластинки определяли с помощью измерительной линейки или штангенциркуля. LS была рассчитана как отношение длины листовой пластинки к её ширине. Площадь листьев была определена весовым методом. Для определения SLA и LDWC части листа (без центральной жилки) известной площади были взвешены для определения свежей массы (FM). Затем высушены до абсолютно-сухой массы (DM) при 80 °С. SLA была рассчитана как отношение площади листа к сухой массе. LWC было рассчитано по формуле:
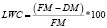 .
.
Для определения содержания фотосинтетических пигментов отбор листьев проводили в 11–12 часов по местному времени. Cодержание фотосинтетичиеских пигментов было определено спектрофотометрическим методом с использованием спектрофотометра (APEL, PD-303, Япония). Содержание хлорофилла a (хл a) и хлорофилла b (хл b) (на сухую массу листа – mass) было определено с использованием уравнений, представленных в работе [10]. Общее содержание хлорофилла (хл общ) было рассчитано как сумма хл a и хл b. Содержание хлорофилла на площадь листа (area) рассчитывали с учетом SLA. Размер светособирающего комплекса (ССК) был рассчитан с предположением, что весь хлорофилл b входит в ССК и соотношение хл a/b в ССК равно 1,2 [5]. Химический анализ был проведен в аналитической лаборатории Института леса КарНЦ РАН. Содержание азота и фосфора определяли спектрофотометрическим методом (спектрофотометр СФ-2000), калия – атомно-эмиссионным методом (атомно-абсорбционный спектрометр AA-7000). Степень фенотипической пластичности (ФП) морфо-биохимических показателей листа была рассчитан по формуле:
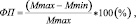
где Mmax – максимальное значение свойства в одном варианте условий;
Mmin – минимальное значение свойства в другом варианте условий [8].
Статистическая обработка данных выполнена с использованием дисперсионного анализа. Достоверность различий средних арифметических оценивали с помощью критерия t Стьюдента (две независимых группы) и критерия Ньюмена-Кейлса (более двух групп). Статистический анализ данных был проведен с использованием программы STATISTICA (StatSoft Inc.) Данные анализировались со степенью надежности 0.95.
Результаты исследования и их обсуждение
Анализ степени пластичности не показал достоверной разницы по усредненному значению пластичности между морфологическими свойствами листа, содержанием фотосинтетических пигментов и элементным составом листа (критерий Ньюмена-Кейлса). Средняя степень пластичности по группам этих признаков составила 13,8; 19,0 и 23,9 % соответственно. Из всех изученных нами показателей листа наибольшей степенью пластичности характеризовалось содержание элементов минерального питания в пересчете на площадь листа, содержание хл a, хл b, хл общ в пересчете на массу листа, отношение хлорофилла к азоту (хл/N), SLA и LA (табл. 1). Показатели, имеющие наибольшую степень пластичности, в основном обеспечивают адаптацию растений к световым условиям [2]. Изменение таких показатели как SLA, содержание питательных веществ и фотосинтетических пигментов, отношение хлорофилла к азоту важно в различных условиях освещенности, так как они связаны с интенсивностью фотосинтеза и балансом углерода в растении [6, 9].
Несмотря на то что свет является важным фактором, влияющим на структурные и функциональными характеристики растений, анализ средней степени пластичности по всем изученным показателям листа у отдельных видов растений показал, что у большинства изученных видов степень пластичности была ниже 20 % (табл. 2).
Низкая степень фенотипической пластичности может быть связана с тем, что высокий уровень пластичности требует больших энергетических затрат, и ее повыщение может быть обратно пропорционально связано со способностью к выживанию в условиях низкой освещенности [7]. Снижение пластичности к световому фактору является частью общей консервативной стратегии использования ресурсов растениями и их высокой устойчивости к затенению [4].
Анализ степени пластичности у разных групп растений, различающихся по месту, которое они занимают в высотном градиенте сообщества, позволил выявить разницу между этими группами в степени пластичности только по LS, содержанию фосфора и содержанию фотосинтетических пигментов (данные не показаны). Степень пластичности по остальным показателям листовой пластинки не отличалась между разными группами. Многими исследователями было показано, что пластичность должна быть большей у видов, которые подвергаются меняющимся условиям освещенности в течение онтогенеза (по мере роста в высоту), по сравнению с видами, которые произрастают постоянно при высокой или низкой освещенности [4]. Мы получили данные подтверждающие большую степень сходства по пластичности исследованных групп растений.
Анализ влияния вида растений и условий освещенности показал, что большая часть изменчивости изученных показателей фотосинтетического аппарата может быть объяснена видовыми различиями (в среднем –79 % вариаций показателей листа). Влияние уровня освещенности было значительно ниже – около 6 %. Несмотря на широкий диапазон варьирования условий освещенности, она оказывала достоверное влияние только на 9 из 24 изученных показателей листа (табл. 3).
Таблица 1
Степень фенотипической пластичности (ФП) показателей листовой пластинки к световому фактору у изученных видов растений
|
Показатели |
ФП, % |
Показатели |
ФП, % |
Показатели |
ФП, % |
|
LL |
12,5±2,1 |
хл общ (mass) |
20,7±3,9 |
ССК |
5,0±0,7 |
|
LW |
9,0±2,2 |
кар (mass) |
11,9±3,2 |
хл/N |
33,1±2,9 |
|
LS |
8,6±1,3 |
хл a (area) |
19,1±2,9 |
N (mass) |
19,3±3,2 |
|
LA |
22,0±3,7 |
хл b (area) |
19,3±3,0 |
P (mass) |
20,7±4,8 |
|
SLA |
20,7±3,4 |
хл общ (area) |
18,8±2,7 |
K (mass) |
16,4±2,8 |
|
LDMC |
9,9±1,4 |
кар (area) |
19,6±2,9 |
N (area) |
29,9±3,3 |
|
хл a (mass) |
20,0±4,0 |
хл a/b |
7,5±1,0 |
P (area) |
28,4±5,4 |
|
хл b (mass) |
22,7±3,8 |
хл/кар |
10,8±2,7 |
K (area) |
28,4±3,5 |
Примечание. Условные обозначения см. в тексте.
Таблица 2
Средняя степень фенотипической пластичности (ФП) показателей листовой пластинки к световому фактору изученных видов растений
|
Вид |
ФП, % |
Вид |
ФП, % |
|
Lagerstroemia calyculata |
19,6 |
Anaxagorea sp. |
12,5 |
|
Dipterocarpus alatus |
21,6 |
Korthalsia sp. |
15,0 |
|
Hibiscus macrophyllus |
21,2 |
Phyllanthus sp. |
18,7 |
|
Dalbergia mammosa |
22,4 |
Ancistrocladus tectorius |
15,0 |
|
Grewia paniculata |
23,9 |
Bauhinia sp. |
19,9 |
|
Pterospermum sp. |
24,2 |
Smilax sp. |
19,8 |
|
Ochrocarpus siamensis |
15,0 |
Korthalsia sp. |
11,1 |
|
Barringtonia cochinchinensis |
15,4 |
Calamus sp. |
21,6 |
Таблица 3
Двуфакторный дисперсионный анализ влияния вида растения и уровня освещенности на морфо-биохимические показатели листовой пластинки
|
Показатели |
Вид |
Освещенность |
Показатели |
Вид |
Освещенность |
||||
|
η2 |
p |
η2 |
p |
η2 |
p |
η2 |
p |
||
|
LL |
96,7 |
*** |
0,4 |
ns |
хл общ (area) |
73,3 |
* |
0,3 |
ns |
|
LW |
97,3 |
*** |
0,4 |
ns |
кар (area) |
75,7 |
* |
0,1 |
ns |
|
LS |
95,7 |
*** |
0,7 |
ns |
хл a/b |
77,6 |
*** |
8,9 |
** |
|
LA |
99,2 |
*** |
0,1 |
ns |
хл/кар |
65,9 |
** |
16,2 |
*** |
|
SLA |
72,5 |
* |
7,6 |
* |
ССК |
65,3 |
* |
0,9 |
ns |
|
LDMC |
94,0 |
*** |
2,4 |
** |
хл/N |
73,9 |
*** |
12,8 |
** |
|
хл a (mass) |
64,9 |
** |
20,0 |
*** |
N (mass) |
90,0 |
*** |
0,9 |
ns |
|
хл b (mass) |
57,2 |
** |
26,0 |
*** |
P (mass) |
84,0 |
** |
0,3 |
ns |
|
хл общ (mass) |
62,1 |
** |
22,9 |
*** |
K (mass) |
94,5 |
*** |
0,01 |
ns |
|
кар (mass) |
77.8 |
*** |
7,8 |
** |
N (area) |
84,1 |
** |
1,2 |
ns |
|
хл a (area) |
73.7 |
* |
0,1 |
ns |
P (area) |
64,2 |
ns |
3,7 |
ns |
|
хл b (area) |
72.7 |
* |
0,6 |
ns |
K (area) |
82,7 |
** |
3,6 |
ns |
Примечание. η2 – сила влияния фактора (эквивалент R2, η2 был рассчитан как отношение факториальной суммы квадратов к общей сумме квадратов *100 %). p – уровень значимости (* p < 0,05; ** p < 0,01; *** p < 0,001, ns – отсутствие достоверного влияния при уровне значимости 0,05).
Таким образом, полученные результаты показали, что каждый из исследованных видов, независимо от места занимаемого им в высотном градиенте сообщества, имеет собственные эндогенные пути адаптации к освещенности. Причем эти пути адаптации различаются, что обеспечивает разный уровень исследуемых морфо-биохимических показателей листа. Возможно, именно эти различия и обеспечивают существование большого видового разнообразия при произрастании в сходных условиях подлеска в биоценозе муссонного тропического леса. Это позволяет предположить, что стратегия адаптации видов в этих условиях направлена на поиски собственного (часто очень узкого) экотопа.
Заключение
Отмеченная для большинства изученных видов низкая степень пластичности, связанная с консервативностью, позволяет растениям занимать определенную узкую стабильную экологическую нишу, снижает риски, связанные с большими энергетическими затратами на поддержание пластичности, и обеспечивает успешное существование растений при низкой доступности ресурсов. Однако эти особенности приводят к тому, что большинство видов растений муссонного тропического леса не способны приспосабливаться к изменениям факторов среды, вызванным естественными и антропогенными нарушениями лесного покрова и связанным, в первую очередь, с увеличением освещенности. Эти особенности, по-видимому, являются одной из системных составляющих, обеспечивающих успешность функционирования этих лесов и их сильную уязвимость к антропогенным нарушениям.
Авторы выражают благодарность руководству Российско-Вьетнамского Тропического центра и администрации национального парка Кат Тиен за возможность проведения исследований.
Библиографическая ссылка
Новичонок Е.В., Марковская Е.Ф., Новичонок А.О. НИЗКАЯ СТЕПЕНЬ ФЕНОТИПИЧЕСКОЙ ПЛАСТИЧНОСТИ РАСТЕНИЙ КАК ОДИН ИЗ ФАКТОРОВ ОРГАНИЗАЦИИ СТАБИЛЬНОЙ СТРУКТУРЫ ТРОПИЧЕСКИХ МУССОННЫХ ЛЕСОВ // Международный журнал прикладных и фундаментальных исследований. 2015. № 7-1. С. 68-72;URL: https://applied-research.ru/ru/article/view?id=6963 (дата обращения: 20.06.2026).